







What is Retina? Structure, Layers and Function of Retina Explained
The retina is the light-sensitive inner layer of the eye that plays a central role in vision. It is located at the back of the eyeball and works as the part of the eye that receives light and converts it into nerve signals. These nerve signals then travel to the brain through the optic nerve, where they are interpreted as images. In simple words, if the eye captures light, the retina helps turn that light into meaningful vision.
Also Check: Structure of Human Eye
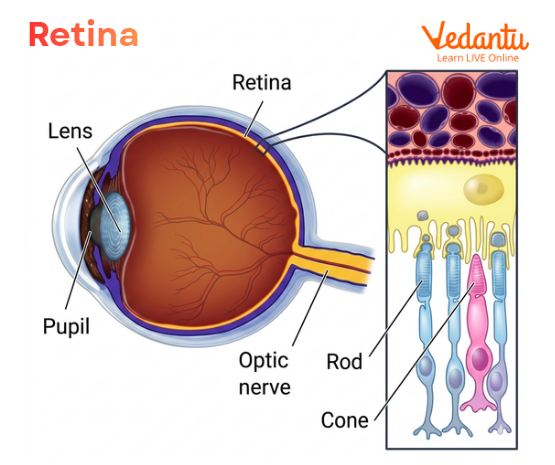
Retina Eye Location
The retina is present at the back of the eye, behind the iris and lens. When light enters the eye and is focused correctly, the rays converge on the retina. This precise focusing is important because the retina must receive a clear image for proper vision.
So, in simple terms:
cornea allows light to enter
lens helps focus it
retina receives that focused light
optic nerve carries the message to the brain
Structure of Retina
The structure of the retina is highly organised and is made of different types of nerve cells arranged in layers. It is not a single flat sheet of identical cells. Instead, it contains photoreceptors, supporting cells, and neural cells that work together to process visual information.
At a basic level, the retina contains:
photoreceptor cells
bipolar cells
ganglion cells
nerve fibre layer
supporting and pigment cells
The retina is adapted to detect light and begin the process of converting it into a nerve impulse. This conversion is called phototransduction. The arrangement of retinal cells allows light detection, signal processing, and onward transmission to the brain.
Parts of the Retina
The retina has two major functional regions:
1. Macula
The macula is the central part of the retina that is responsible for sharp central vision. It is especially important for seeing colours and fine details. This helps us perform activities such as:
reading
recognising faces
driving
seeing fine patterns and small objects
The cones are concentrated in the macular region, which is why this area is associated with detailed and colour vision.
2. Peripheral Retina
The peripheral retina helps us see objects to the side while looking straight ahead. It is more important for side vision and for seeing in dim light. Rod cells are more important in this region. These rods help detect movement and function well under low-light conditions.
This is why damage to the peripheral retina may cause narrowing of vision or tunnel vision.
Function of Retina
The main function of the retina is to detect light and convert it into nerve signals. These signals are then sent to the brain, where they are interpreted into meaningful visual images.
Detailed Functions of Retina
1. Light Detection
The retina contains photoreceptor cells that react to light. These cells absorb light and initiate a neural response.
2. Conversion of Light Into Nerve Signals
This is the central physiological role of the retina. Light energy is changed into electrical impulses through photoreceptor activity.
3. Transmission to the Brain
The signals generated in the retina are carried through the optic nerve to the visual areas of the brain.
4. Central and Peripheral Vision
The retina helps in both central detailed vision and peripheral side vision, depending on the region involved.
5. Colour Vision and Low-Light Vision
Different photoreceptors in the retina support different visual tasks. Cones are mainly for colour and detail, while rods are mainly for dim light and peripheral vision.
This is why retinal damage can create different kinds of visual loss depending on which retinal area is affected. Some people may lose central vision, some may lose side vision, and some may develop blind spots.
Photoreceptors in the Retina
Photoreceptors are the light-sensitive cells of the retina. They are essential for converting light into visual signals.
There are two main types of photoreceptors:
Rods
Rods are more sensitive to dim light. They help in:
night vision
peripheral vision
motion detection
Rods do not help much in colour vision.
Cones
Cones function best in bright light. They help in:
colour vision
sharp central vision
fine detail recognition
The cones are highly concentrated in the macula. This is why the macula is so important for reading and recognising faces.
Layers of Retina
The layers of the retina are an important part of eye anatomy. The retina is made of several distinct layers of cells and fibres arranged in a specific order. These layers help receive light, process visual signals, and pass them to the optic nerve.
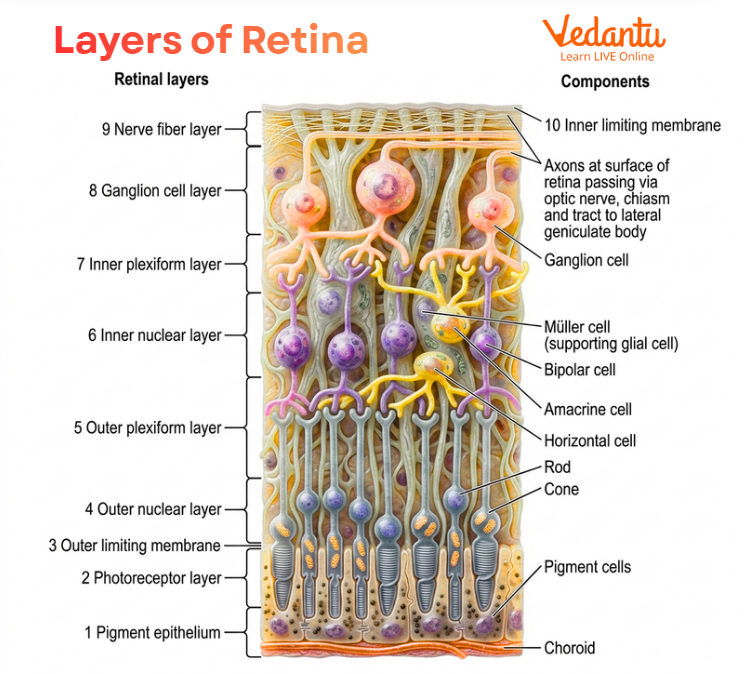
Retinal pigment epithelium
Layer of rods and cones
External limiting membrane
Outer nuclear layer
Outer plexiform layer
Inner nuclear layer
Inner plexiform layer
Ganglion cell layer
Nerve fibre layer
Internal limiting membrane
1. Retinal Pigment Epithelium
This is the outermost retinal layer. It supports photoreceptors and helps absorb excess light, preventing internal reflection.
2. Layer of Rods and Cones
This contains the outer and inner segments of photoreceptor cells. It is the actual light-detecting layer.
3. External Limiting Membrane
This is a thin boundary-like layer separating photoreceptor segments from their nuclei.
4. Outer Nuclear Layer
This contains the nuclei of rods and cones.
5. Outer Plexiform Layer
This is the region where photoreceptors form synapses with bipolar and horizontal cells.
6. Inner Nuclear Layer
This layer contains the nuclei of bipolar cells, horizontal cells, amacrine cells, and Müller supporting cells.
7. Inner Plexiform Layer
This is where bipolar cells connect with ganglion cells and other interneurons.
8. Ganglion Cell Layer
This contains the cell bodies of ganglion cells.
9. Nerve Fibre Layer
This consists of the axons of ganglion cells, which converge to form the optic nerve.
10. Internal Limiting Membrane
This is the innermost retinal layer facing the vitreous humour.
Retina Problems and Diseases
Many retina problems can affect vision seriously. Since the retina is responsible for converting light into nerve signals, damage to it can produce permanent vision loss if not treated in time.
Common retina problems include:
age-related macular degeneration
diabetes-related retinopathy
hypertensive retinopathy
macular hole
macular pucker
ocular migraine
posterior vitreous detachment
retinal bleeding
retinal detachment
retinal tears
retinal vein occlusion
retinal artery occlusion
retinal inflammation or uveitis
retinitis pigmentosa
retinopathy of prematurity
solar retinopathy
eye cancers such as retinoblastoma
colour blindness including achromatopsia
Symptoms of Retina Problems
If the retina is damaged or diseased, a person may develop noticeable visual symptoms. Common signs of retina problems include:
blurry vision
distorted vision
peripheral vision loss or tunnel vision
double vision
flashes of light
floaters
light sensitivity
blind spots
worsening vision over time
Sudden changes in vision are especially serious and should not be ignored because some retinal conditions can progress rapidly and lead to irreversible blindness if not treated promptly.
Tests Used to Examine the Retina
An eye specialist checks the retina during a complete eye examination. To see the retina clearly, the pupil is often dilated.
Important retinal examination methods include:
1. Fundoscopy or Ophthalmoscopy
This helps directly examine the inside of the eye, including the retina.
2. Slit Lamp Examination
This gives a magnified view of the eye and can be used along with special lenses to study retinal structures.
3. Fundus Photography
This provides retinal images for documentation and comparison.
4. Red Reflex or Fundus Reflex Testing
This helps detect abnormalities in the eye.
5. Visual Acuity Testing
This measures how clearly a person can see.
6. Visual Field Testing
This checks central and peripheral vision.
These tests are especially important in people with diabetes, hypertension, or progressive visual symptoms.
Retina Care and Prevention
Protecting the retina is very important for preserving vision. Good retinal care includes both routine eye examination and daily precautions.
get regular eye examinations every one to two years
if you have diabetes or other chronic conditions, get eye checks at least once a year
maintain a healthy body weight
focus on proper nutrition
wear protective eyewear during sports or risky activities
use sun protection such as good quality sunglasses
avoid exposure to extremely bright light sources
do not ignore even gradual vision changes
Prompt medical care is very important if sudden vision loss or sudden flashes, floaters, or blind spots appear.
Why the Retina is Important in Vision?
The retina is important because it is the structure that begins the entire process of vision. Without the retina, incoming light would not be converted into neural information. Even if the cornea and lens worked perfectly, vision would still not occur without a functioning retina.
Why the retina matters so much:
it detects light
it distinguishes colour and brightness
it supports central and peripheral vision
it starts the transmission of visual messages
it connects the eye to the brain through the optic nerve
This is why the retina is often called the sensory screen of the eye, though in reality it does far more than a simple screen.
FAQs on Retina: Structure, Layers, Functions, Diagram and Retina Problems for NEET
1. What is the retina and its function?
The retina is a thin, light-sensitive layer at the back of the eye. Its main function is to detect light and convert it into nerve signals, which are then sent to the brain through the optic nerve for vision.
2. What are signs of retina problems?
Common signs of retina problems include:
Sudden floaters (small spots or threads)
Flashes of light
Blurred or distorted vision
Loss of side (peripheral) vision
A dark shadow or curtain over vision
3. Can retina damage be repaired?
Retina damage can often be treated or controlled with medical procedures like laser therapy or surgery. However, early treatment is important, as damaged retinal cells do not fully regenerate, and vision loss may not be completely reversible.
4. Why is it called retina?
The name retina comes from the Latin word rete, meaning “net”, because it has a network-like structure of nerve cells.
5. What are the 5 diseases of the retina?
Five common retina diseases are:
Diabetic retinopathy
Macular degeneration
Retinitis pigmentosa
Retinal vein occlusion
Retinal tear or detachment
6. What are the first signs of a retinal tear?
Early signs of a retinal tear include:
Flashes of light
Sudden increase in floaters
Shadow or curtain over part of vision
Blurred or reduced vision
These symptoms need immediate medical attention.