

What are plastids definition structure types and functions
Imagine a bustling factory within every plant cell where specialised workers convert sunlight into energy and store vital nutrients. Plastids, the double-membrane organelles found in plant cells, serve as these essential factories. From photosynthesis in chloroplasts to pigment production in chromoplasts, plastids play diverse roles that keep plants thriving. In this engaging guide, we explore what are plastids, their structure, various types, and their functions in plant cells. With surprising facts and interactive elements, you'll gain a deeper understanding of these fascinating organelles and their impact on both plant biology and everyday life.
Understanding Plastids: Definition and Function
Plastids definition: Plastids are specialised, double-membrane organelles in plant cells responsible for manufacturing and storing food, pigments, and other vital compounds. They are crucial for processes like photosynthesis and nutrient storage, making them indispensable for plant survival.
What is plastids?
Plastids are organelles that contribute to key cellular functions, such as converting sunlight to energy (in chloroplasts) and storing compounds (in leucoplasts).
Function of plastids:
Photosynthesis: Chloroplasts capture light energy and transform it into chemical energy.
Pigment Storage: Chromoplasts store pigments that lend leaves, flowers, and fruits their vibrant colours.
Nutrient Storage: Leucoplasts store starches, proteins, and lipids, ensuring plants have the energy and building blocks they need.
Types of Plastids and Their Specialisations
Types of plastids vary based on the presence of pigments and their developmental stages. The main types include:
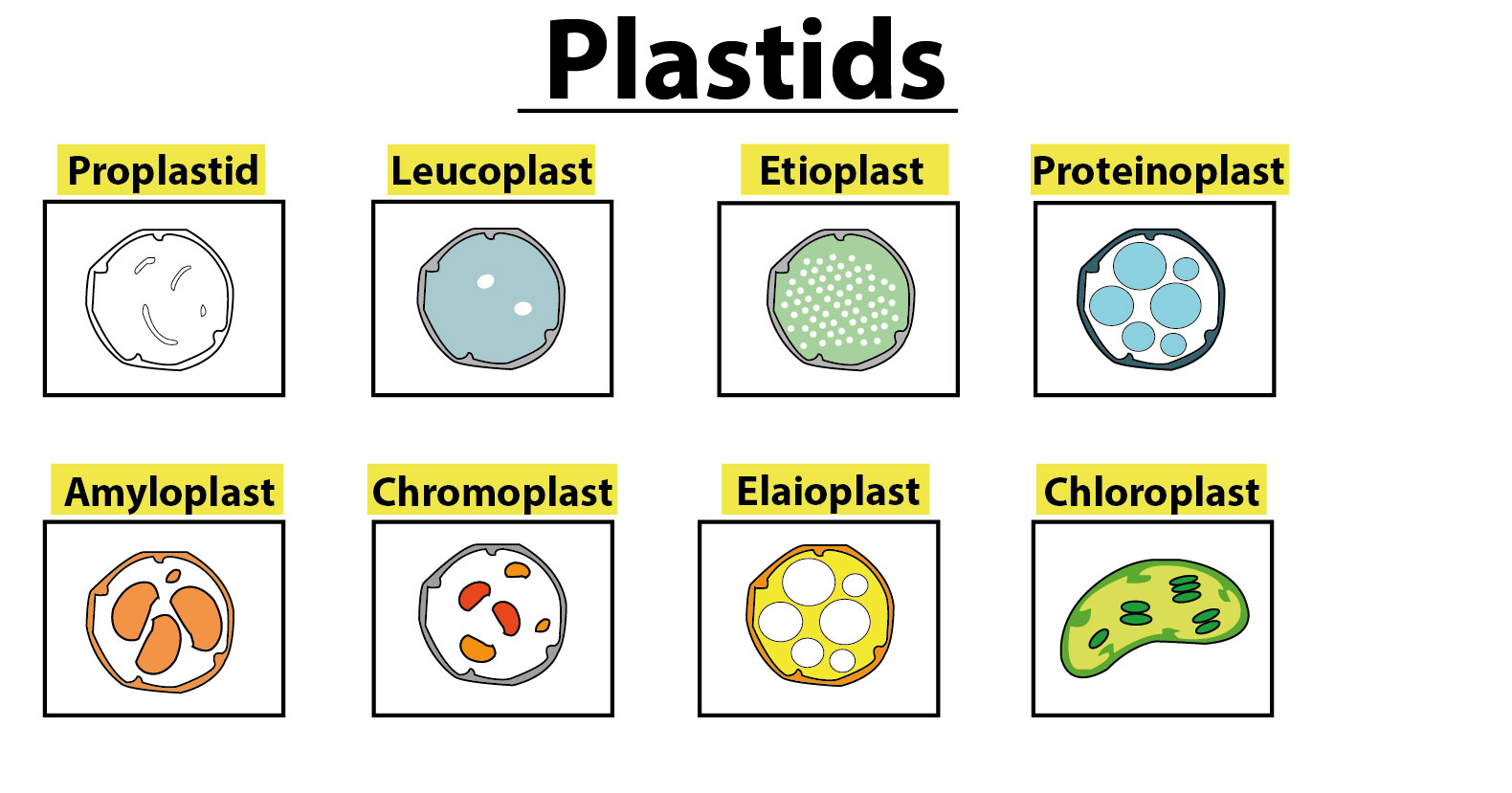
Chloroplasts
Structure of plastids: Chloroplasts are biconvex, semi-porous, double-membrane organelles located mainly in the mesophyll of plant cells.
Role: They are the primary sites for photosynthesis, converting light energy into sugars.
Chromoplasts
Plastids in plant cells: Chromoplasts, found in flowers, ageing leaves, and fruits, store carotenoid pigments.
Significance: They provide the bright colours necessary for attracting pollinators and enhancing visual appeal.
Gerontoplasts
Transition Role: Gerontoplasts are ageing chloroplasts that transition as the leaf's photosynthetic activity declines, especially during autumn.
Leucoplasts
Non-Pigmented Organelles: These are colourless plastids found in non-photosynthetic tissues like roots.
Subtypes:
Amyloplasts: Specialised in storing and synthesising starch.
Proteinoplasts: Involved in the storage of proteins, often found in seeds.
Elaioplasts: Dedicated to the storage of fats and oils.
Students can also read out detailed article on the Difference between mitochondria and plastids to understand the concept better.
- Inheritance and the Uniparental Trait
Plastids are inherited uniparentally. In angiosperms, plastids are passed on through the female gamete, while in many gymnosperms, the male pollen is the source. This unique mode of inheritance underscores the critical role of plastids in plant development and evolution.
Test your Knowledge on Plastids!
Take this short quiz and then “Check your answers” to see how well you understand plastids.
Question: What are plastids primarily responsible for in plant cells?
Question: Name the type of plastid that is essential for photosynthesis.
Question: Which plastid is known for storing pigments that give fruits and flowers their colour?
Question: What is the function of leucoplasts in non-photosynthetic parts of the plant?
Question: How are plastids generally inherited in angiosperms?
Check Your Answers:
Manufacturing and storing food, pigments, and other compounds.
Chloroplasts.
Chromoplasts.
They store starch, proteins, or lipids.
Through the female gamete (uniparental inheritance).
Fun Facts about Plastids
Fact 1: Despite being small, plastids can transform into different forms (e.g., chloroplasts converting into chromoplasts) as plants mature.
Fact 2: Plastids contain their own DNA, highlighting their evolutionary history as once free-living organisms.
Fact 3: The vibrant colours of autumn leaves are partly due to the transformation of chloroplasts into gerontoplasts.
Real-World Applications
Plastids are not just academic curiosities—they have real-world significance:
Agriculture: Understanding plastids helps in developing crops with enhanced nutritional qualities and improved stress resistance.
Biotechnology: Manipulating plastid functions is a key strategy in producing biofuels and biopharmaceuticals.
Horticulture: The role of chromoplasts in colour production is exploited to breed visually appealing fruits and ornamental plants.


FAQs on Plastids in Plant Cells Structure Types and Roles
1. What are plastids in plant cells?
Plastids are membrane-bound organelles found in plant cells and algae that are responsible for photosynthesis, storage, and pigment synthesis. They are double-membraned structures containing their own DNA and ribosomes, making them semi-autonomous. Plastids develop from immature precursors called proplastids and differentiate into various types depending on cellular needs.
2. What are the main types of plastids?
The three main types of plastids are chloroplasts, chromoplasts, and leucoplasts.
- Chloroplasts – Carry out photosynthesis and contain chlorophyll.
- Chromoplasts – Store carotenoid pigments that give fruits and flowers their color.
- Leucoplasts – Colorless plastids mainly involved in storage of starch, oils, or proteins.
3. What is the function of chloroplasts?
The main function of chloroplasts is to perform photosynthesis, converting light energy into chemical energy in the form of glucose. This process occurs in two stages:
- Light reactions – Occur in the thylakoid membranes and produce ATP and NADPH.
- Calvin cycle – Occurs in the stroma and synthesizes glucose from carbon dioxide.
4. What is the difference between chloroplasts and chromoplasts?
The main difference between chloroplasts and chromoplasts is that chloroplasts perform photosynthesis, while chromoplasts store pigments.
- Chloroplasts contain chlorophyll and are green in color.
- Chromoplasts contain carotenoids and provide red, orange, or yellow colors to fruits and flowers.
5. What are leucoplasts and what do they do?
Leucoplasts are colorless plastids that primarily function in the storage of food materials. They are commonly found in non-photosynthetic tissues like roots and seeds. Types of leucoplasts include:
- Amyloplasts – Store starch.
- Elaioplasts – Store oils and lipids.
- Proteinoplasts – Store proteins.
6. Do plastids have their own DNA?
Yes, plastids contain their own circular plastid DNA (ptDNA), which allows them to synthesize some of their own proteins. Plastid DNA resembles bacterial DNA, supporting the endosymbiotic theory. However, many plastid proteins are still encoded by nuclear genes and imported into the plastid.
7. How do plastids develop in plant cells?
Plastids develop from undifferentiated precursor organelles called proplastids found in meristematic cells. Their development depends on environmental and cellular conditions:
- In the presence of light, proplastids develop into chloroplasts.
- In storage tissues, they become leucoplasts.
- During fruit ripening, chloroplasts may convert into chromoplasts.
8. What is the structure of a chloroplast?
A chloroplast has a double membrane and an internal system of thylakoids arranged in stacks called grana. Its main structural components include:
- Outer membrane – Protective covering.
- Inner membrane – Regulates transport.
- Stroma – Fluid matrix where the Calvin cycle occurs.
- Thylakoids – Flattened sacs containing chlorophyll.
9. Why are plastids important for plants?
Plastids are essential because they enable photosynthesis, pigment formation, and food storage in plants. Their importance includes:
- Production of glucose through photosynthesis.
- Synthesis of pigments like chlorophyll and carotenoids.
- Storage of starch, oils, and proteins.
10. Are plastids found in animal cells?
No, plastids are not found in animal cells; they are present only in plants and algae. Animal cells lack chloroplasts and other plastids because they do not perform photosynthesis. Plastids are a defining feature of plant cells and contribute to their autotrophic mode of nutrition.










