

Difference between cell wall and cell membrane with structure and functions
Cells are the fundamental units of life, often described as the building blocks of organisms. Dating back to Robert Hooke’s discovery in 1665, cells come in various shapes and sizes depending on their functions and compositions. In this guide, we will explore the roles of the cell wall and cell membrane, discuss the difference between cell wall and cell membrane, and explain why each is important for the life of a cell.
What is a Cell Wall?
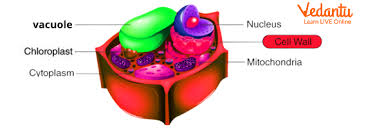
The cell wall is a rigid, outer covering found in plant cells, fungi, some bacteria, and algae. It lies outside the cell membrane and is crucial in providing structural support. Here are the key points:
Composition: In plant cells, the cell wall is primarily made of cellulose, along with components such as hemicellulose, lignin, and pectin. In fungi, chitin is a major component, while bacteria have peptidoglycan.
Functions:
Protection: Safeguards the cell against physical damage and pathogens.
Support: Maintains the shape and structural integrity of the cell.
Growth Regulation: Controls the direction and extent of cell growth.
Storage: Acts as a storage area for carbohydrates, particularly in seeds.
Permeability: Allows the free passage of small molecules.
Unlike the cell membrane, the cell wall not only provides mechanical strength but also plays a role in intercellular communication by transmitting signals that can influence growth and development. This aspect is particularly vital during plant responses to environmental stress.
Also, read Plant Cell
What is a Cell Membrane?
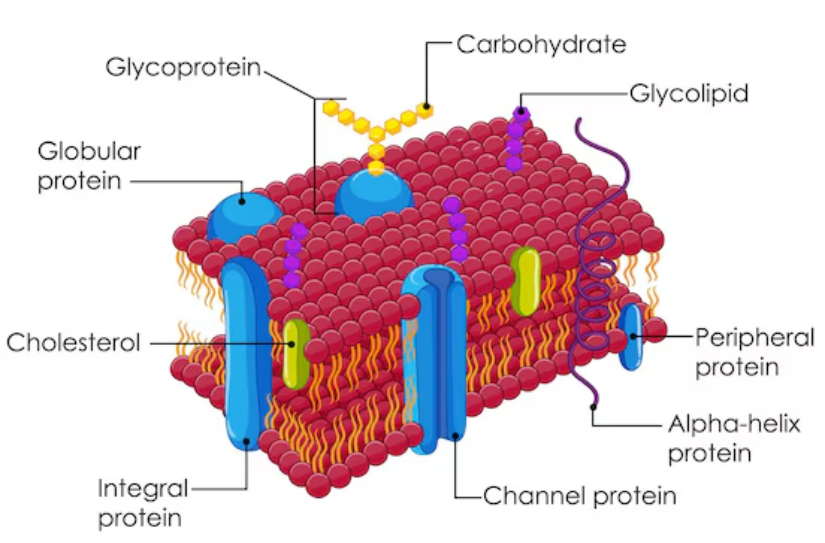
The cell membrane (or plasma membrane) is a thin, semi-permeable barrier found in all cells, including those of plants, animals, bacteria, and fungi. It is essential for maintaining the internal environment of the cell. Key features include:
Composition: The cell membrane is primarily a lipid bilayer interspersed with proteins and carbohydrates.
Functions:
Selective Permeability: Regulates the movement of substances into and out of the cell, ensuring only selected molecules pass through.
Protection: Acts as a barrier protecting the cell’s internal components.
Communication and Signalling: Contains receptors that help in cell-to-cell communication.
Support and Structure: Maintains the cell’s shape and aids in processes such as cell division.
Metabolic Activity: Unlike the cell wall, the cell membrane is metabolically active and adapts to changes in the cell’s environment.
The function of cell membranes is central to maintaining homeostasis, and understanding the importance of cell membranes is crucial in biology.
Also, read Cell membrane
Difference Between Cell Wall and Cell Membrane
It is common to ask, "is cell wall and cell membrane the same?" The answer is no. They are distinct structures with unique roles:
Understanding the difference between cell wall and cell membrane is vital for recognising how cells maintain structure and regulate internal processes.
Explore more: Difference between Plant Cell and Animal Cell
Real-World Applications
The study of cell walls and cell membranes is not just academic—it has many practical applications:
Medicine and Pharmacology: Knowledge of the function of cell membrane is crucial in drug delivery systems, where targeting cell receptors improves therapeutic outcomes.
Agriculture: Understanding cell walls helps in developing crops that are more resistant to pests and environmental stresses.
Biotechnology: Engineers use cell membrane properties in the design of biosensors and in the development of artificial cells.
Food Industry: The manipulation of cell wall components can improve food texture and shelf life.
These applications highlight the importance of cell membrane and the role of the cell wall in various industries, making this topic highly relevant in real-life scenarios.
Fun Facts about Cell Walls and Cell Membranes
Historical Insight: The term "cell" was coined by Robert Hooke in 1665 after observing the tiny compartments in cork, which resembled the small rooms of a monastery.
Adaptive Structures: Some bacteria can alter their cell wall composition to resist antibiotics, showcasing nature’s ingenuity.
Dynamic Membranes: Despite its thin structure, the cell membrane is incredibly dynamic, continuously changing its composition in response to environmental signals.
Key Points
Intercellular Communication: Recent studies reveal that the cell wall is not merely a passive barrier but can actively participate in cell signalling, impacting how cells react to their surroundings.
Membrane Fluidity: The fluid nature of the cell membrane allows cells to merge with one another during processes such as fertilisation and immune responses.
Evolutionary Perspective: The evolution of cell walls and membranes has been central to the development of complex life forms, influencing the diversity of life on Earth.


FAQs on Cell Wall and Cell Membrane Explained for Students
1. What is the difference between a cell wall and a cell membrane?
The main difference between a cell wall and a cell membrane is that the cell wall is a rigid outer layer for protection and support, while the cell membrane is a flexible, selectively permeable barrier that controls movement of substances.
- Cell wall: Present in plants, fungi, bacteria, and algae; made of cellulose (plants), chitin (fungi), or peptidoglycan (bacteria); provides shape and mechanical strength.
- Cell membrane: Present in all cells; made of a phospholipid bilayer with proteins; regulates transport and communication.
- The cell wall lies outside the cell membrane in organisms that have both.
2. What is the function of the cell wall?
The primary function of the cell wall is to provide structural support and protection to the cell.
- Maintains the cell’s shape.
- Prevents bursting due to osmotic pressure in hypotonic environments.
- Offers protection against mechanical injury and pathogens.
- Helps maintain turgor pressure in plant cells.
3. What is the function of the cell membrane?
The main function of the cell membrane is to regulate the movement of substances into and out of the cell.
- Acts as a selectively permeable membrane.
- Controls transport through diffusion, osmosis, and active transport.
- Contains receptor proteins for cell signaling.
- Maintains the cell’s internal environment or homeostasis.
4. What is the cell membrane made of?
The cell membrane is made of a phospholipid bilayer with embedded proteins, carbohydrates, and cholesterol.
- Phospholipids: Form a double layer with hydrophilic heads and hydrophobic tails.
- Membrane proteins: Help in transport and communication.
- Cholesterol: Maintains membrane fluidity (in animal cells).
- Carbohydrates: Form glycoproteins and glycolipids for cell recognition.
5. What is the cell wall made of in plants?
The plant cell wall is mainly made of cellulose, a structural polysaccharide.
- Cellulose fibers provide tensile strength.
- Embedded in a matrix of hemicellulose and pectin.
- Some plant cells also develop a secondary wall containing lignin for extra rigidity.
6. Is the cell membrane present in plant and animal cells?
Yes, the cell membrane is present in both plant and animal cells.
- In animal cells, it is the outermost boundary.
- In plant cells, it lies just inside the cell wall.
- It performs the same essential functions of selective transport and communication in both types.
7. Why do plant cells have a cell wall but animal cells do not?
Plant cells have a cell wall to provide rigidity and support, while animal cells lack it to allow flexibility and movement.
- Plants need structural strength to stand upright.
- The cell wall helps maintain turgidity in plant tissues.
- Animal cells require flexibility for processes like movement and phagocytosis.
8. What is selectively permeable in the cell membrane?
The cell membrane is selectively permeable, meaning it allows some substances to pass while restricting others.
- Small nonpolar molecules like oxygen diffuse freely.
- Water moves by osmosis.
- Ions and large molecules require channel proteins or carrier proteins.
- This selectivity helps maintain cell homeostasis.
9. How does the cell membrane control the movement of substances?
The cell membrane controls movement through passive and active transport mechanisms.
- Passive transport: Includes diffusion and osmosis; does not require energy.
- Facilitated diffusion: Uses transport proteins without energy.
- Active transport: Requires ATP to move substances against a concentration gradient.
- Endocytosis and exocytosis: Move large particles in vesicles.
10. What is the fluid mosaic model of the cell membrane?
The fluid mosaic model describes the cell membrane as a flexible layer of lipids with proteins embedded within it.
- “Fluid” refers to the lateral movement of phospholipids.
- “Mosaic” refers to the patchwork arrangement of proteins.
- Proposed by Singer and Nicolson (1972).
- Explains membrane flexibility and selective permeability.










