





Revision Notes on Sexual Reproduction in Flowering Plants for NEET 2024 - Free PDF Download
Introduction
The reproduction process occurs in the plant through a special structure known as the flower.
The typical or complete flower has a whorl of sepals (calyx), a whorl of petals (corolla), androecium (male part) of stamens, and gynoecium (female part) of carpels.
Because the calyx and corolla do not engage in the reproduction process, they are referred to as accessory whorls of the flower.
However, the androecium and gynoecium are directly related to reproduction, and thus, are known as essential whorls.
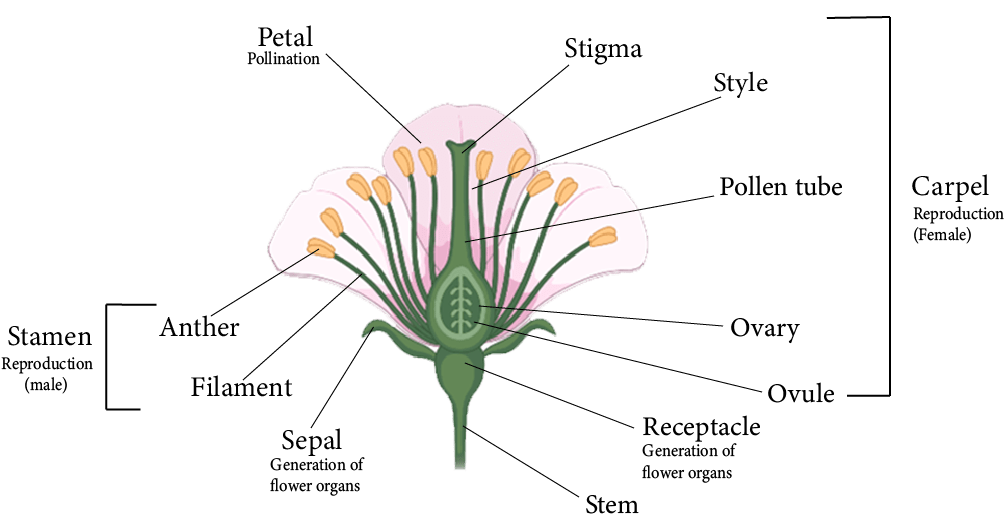
Parts of a Flower
Sexual Reproduction
Sexual reproduction is the procedure by which new organisms are created through the formation and fusion of gametes.
Stamens are male reproductive organs in flowering plants, while carpels or pistils are female reproductive organs.
Meiosis is the process by which male and female gametes are created respectively in male and female sex parts in angiosperms.
Both gametes fuse to establish a diploid zygote, which leads to embryo formation. This is known as fertilisation.
The process of growth and development of androecium (male reproductive organ) and gynoecium (female reproductive organ), pollination, and fertilisation are all pre-fertilisation events.
Endosperm development, embryo development (in dicots and monocots), and seed development are all post-fertilisation events.
Male Reproductive Parts
The male reproductive organ is known as the androecium, and its reproductive unit is known as the stamen.
Microsporophyll is another name for the stamen.
A typical stamen divides into three parts:
A long, thin structure called the filament connects the stamen to the flower's thalamus.
The anther is a swollen spore-bearing structure at the free end of the filament.
An anther and a filament are joined together by a small region known as a connective. Connective incorporates vascular tissues.
Each anther is a bilobed structure, meaning it has two anther lobes and is termed dithecous.
Each anther lobe has two chambers known as pollen sacs or pollen chambers.
As a result, the typical anther contains four pollen sacs, thus, such anther is called tetrasporangiate.
Pollen grains are formed within the pollen sac via meiotic division of the pollen mother cells.
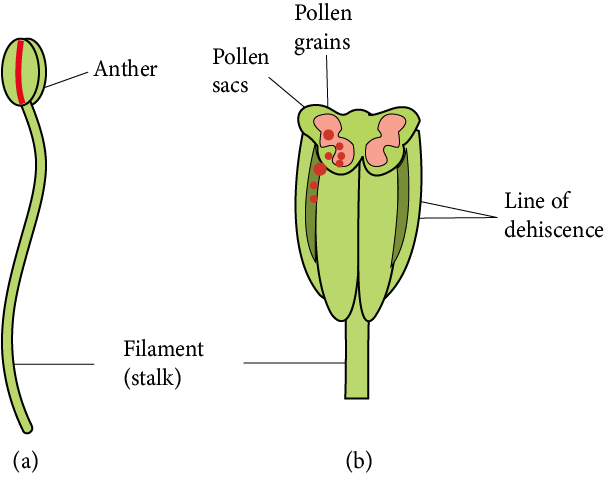
Male Reproductive Parts
Structure of Anther:
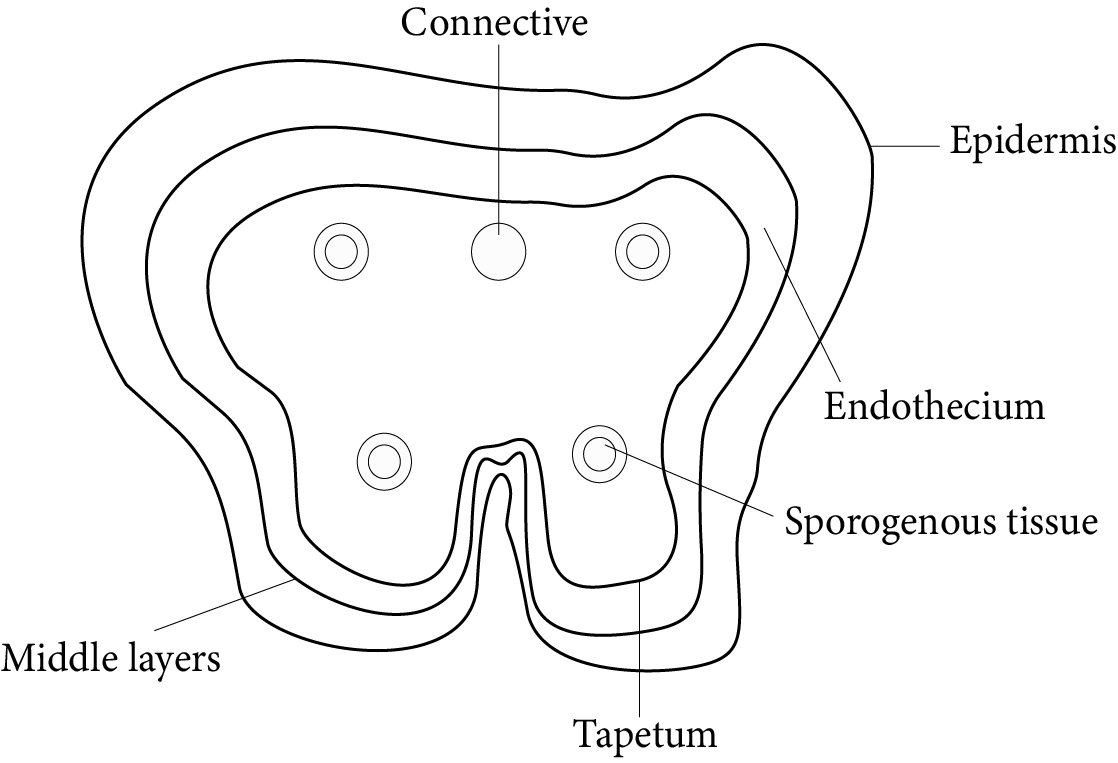
Lateral Section of Anther
The origin of the anther is Eusporangiate, which means that it develops from more than one archesporial cell.
It appears almost spherical when looked at in its transverse section.
The wall layers of the anther are as follows: epidermis, endothecium, middle layer, and tapetum.
Epidermis
It is the anther's outermost protective layer which is single-celled and continuous.
Endothecium
This layer exists beneath the epidermis. It is a thick single-celled layer.
Various changes occur in the walls of endothecium cells during the development of the anther.
Because of the availability of fibrous thickening, the endothecium becomes hygroscopic.
As a result, it aids in the dehiscence of the anther.
Middle layer
The middle layer is made up of parenchymatous cells.
It has one to three celled thicknesses and its parenchymatous cells store food.
Tapetum
Tapetum is the nutritive layer which is the single-celled and present innermost inside the anther.
It surrounds the pollen sacs.
The cells of the tapetum layer are at first diploid, but due to endomitosis, they become polyploid.
Tapetum receives food from the middle layer and feeds microspore mother cells or microspores present in the pollen sacs.
Tapetum cells naturally produce hormones and callase enzymes. In the mature anther, the tapetum layer disappears.
The tapetal cells contribute significantly during the meiotic cell division of microspore mother cells and in pollen development.
Microsporogenesis:
Microsporogenesis is the process of the creation of microspores or pollen grains inside the anther.
An anther in its mature state is a clump of undifferentiated and homologous meristematic cells surrounded by the epidermis.
First, vascular tissue forms in the middle region. Four cells just beneath the epidermis grow in size at the same time.
These are the Archesporial cells with a visible nucleus and dense cytoplasm.
Archesporial cells divide periclinal to produce primary parietal cells beneath the epidermis and primary sporogenous cells in the centre.
Primary parietal cells divide further periclinal and anticlinal to form a series of 3-5 layers that form the anther walls.
Primary sporogenous cells split twice or more than twice by mitotic division to form sporogenous cells, which later differentiate into microspore mother cells (MMC) during pollen sac wall formation.
By meiotic or reduction division, each MMC divides into four haploid microspores or pollen grains.
In the beginning, all four microspores are joined together by a callose layer. Tetrad is the name given to this group of microspores. This callose layer is eventually dissolved by the callase enzyme, which is secreted by the tapetum.
Tetrads are of various types based on the arrangement of microspores in them. These types include:
Tetrahedral tetrad (most common)
Isobilateral tetrad
Decussate tetrad
T-Shaped tetrad
Linear tetrad
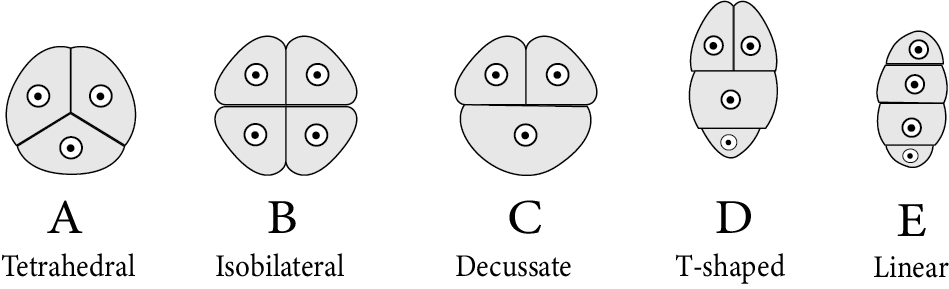
Types of Microspore Tetrads
Structure of Pollen Grain or Microspore:
A pollen grain is a male gametophyte's first cell and is enclosed by two distinct layers.
The exine is the thick, rigid, and ornamented outer layer (wall). Cutin and sporopollenin combine to form this layer. Sporopollenin is a very tough material and it does not biodegrade. It creates distinctive sculpturing or designs on the surface of pollen grains.
The inner layer is thin, soft, and pliable. It's known as intine and is composed of pectin and cellulose (or pecto-cellulose).
The outer surface exine is generally absent at a few locations on pollen grain. These thin areas are known as germ pores.
During pollen grain germination, the intine emerges from any one germ pore as a pollen tube.
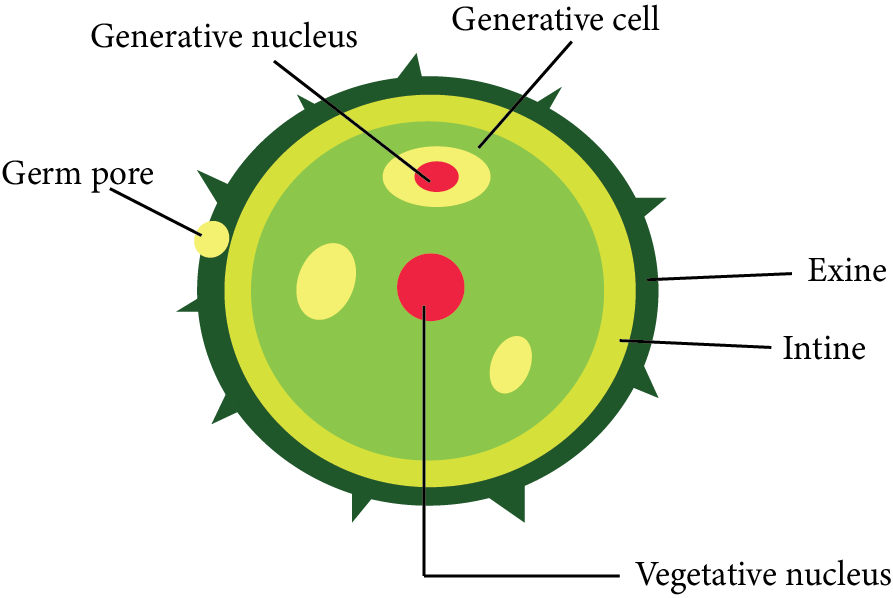
Structure of Pollen Grain
Pollen grains from plants that are pollinated by insects have an oily, thick, and sticky layer around them. It's called a pollen kit, and it's made up of lipids and carotenoids.
The pollen kit's functions are as follows:
This oily layer provides protection to the pollen grain from harmful ultraviolet rays.
Its sticky surface aids in attachment to the insects.
The yellow colour attracts insects.
All pollen grains in the plants belonging to Asclepiadaceae and Orchidaceae families join together to form "Pollinium."
Pollen grains from certain plants present in the air that cause allergy are known as "aeroallergens," such as Parthenium, Sorghum, Chenopodium, and Amaranthus.
Pollen from Ambrosia causes "hay fever."
Zostera, an aquatic plant, has the longest pollen.
Dehiscence of Anther:
Various changes occur in the walls of the anther during maturation.
Initially, the middle layer degrades due to food absorption by the tapetum. Tapetum degenerates over time as well.
The tapetum cells contain numerous spherical bodies named pro-ubisch bodies only when anther is young, at the sporogenous tissue phase. Meiosis increases the amount of pro-ubisch bodies in pollen mother cells. These pro-ubisch bodies become ubisch bodies or orbicules after being coated by sporopollenin.
As a result, the outer covering of a mature anther consists of only two layers: epidermis and endothecium.
Anther lobes degenerate too in a mature anther. As a result, the pollen sacs of every anther lobe transform into a single pollen sac. As a result, only two pollen sacs are available in a mature anther.
Anther dehiscence occurs during the dry season. Because of the hygroscopic nature of endothecium, water is lost from the cell.
Endothelial cell walls try to contract due to water loss and create a concave shape or become incurved.
The curvature of the outer walls exerts a pulling force or tension across the exposed surface of the anther.
Tension causes thin-walled stromal cells to break off, dehiscence of the anther, and pollen grains to be present in pollen sacs dispersed into the atmosphere.
Anther dehiscence in angiosperms can occur either by longitudinal, apical pore, transverse or valvular mode.
Development of Male Gametophytes:
Pollen grain (microspore) is the first cell of a male gametophyte in flowering plants.
A generative cell splits by mitosis inside the pollen tube to form two nonmotile male gametes.
Male gametophytes now have a three-celled structure with one vegetative cell as well as two male gametes.
This three-celled phase describes an angiosperm's mature male gametophyte.
Male gametophyte is severely reduced and entirely dependent on sporophyte.
Development of Pollen Grain During Pre-pollination:
At the start of the process, the unequal mitotic division only divides the nucleus of the pollen grain into two nuclei of unequal size.
A small nucleus near the wall is known as a generative nucleus, and a large nucleus inside the cytoplasm is known as a tube or vegetative nucleus.
Both the nucleus and the cytoplasm surrounding it become dense, which is pursued by unequal cytokinesis, leading to the formation of two cells of unequal size.
A vegetative cell is a larger cell with a large nucleus, whereas a generative cell is a smaller cell with a small nucleus.
This pollen grain stage is known as a partially developed male gametophyte.
Development of Pollen Grain After Pollination:
After pollination, the remainder of the pollen grain development [Immature male gametophyte] occurs on the stigma of the carpel.
The moisture and sugar composition of the stigma are absorbed by pollens.
The volume of internal cytoplasm grows as a result of this. It applies pressure to both inner and outer layers, intine and exine. Because of this pressure, intine emerges from any germ pore in the shape of a tube-like structure known as a pollen tube.
The vegetative nucleus first enters the pollen tube and reaches the terminal position.
This spindly generative cell has now entered the pollen tube.

Development of Pollen Grain After Pollination
Female Reproductive Parts
In angiosperms, the flowers possess the female reproductive body known as gynoecium.
Megasporophyll is another name for carpel.
The carpel is classified into three distinct regions:
Stigma
Style
Ovary
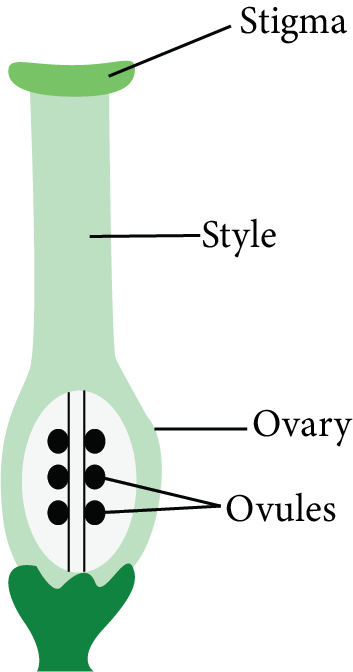
Structure of Carpel
The stigma is the free end or receptacle of the carpel that receives pollen grains.
The style is a long, slender tubular structure that exists between the stigma and the ovary and joins them.
The ovary is the bulging basal part of the carpel. The ovules (megasporangium) are attached to the ovary wall on a cushion-like cell mass named the placenta.
Inside the ovary, one or more ovules are available.
Structure of Ovule or Megasporangium
The placenta inside the ovary forms a ridge or stalk-like subsequent growth on which the body of ovules is present.
Each ovule is connected to the placenta by a thin stalk known as a funicle or funiculus or funiculus.
The hilum is the point at which the funicle connects to the ovule.
The nucellus is a volume of parenchymatous cells that makes up the main zone of the ovule. The nucellus is the most important part of the ovule.
The nucellus is surrounded by one or two coats known as integuments.
Chalaza is the location where funicles and integuments emerge. It is present at the opposite end of the micropyle.
Because a small site opposite the chalaza lacks the integuments, a tiny gap (pore) forms, which is known as a micropyle.
Some angiosperms retain a portion of the nucellus within the ovules. Perisperm is that retained part of the nucellus found inside the seed as a thin layer. It is found in the plants of families Piperaceae (Piper nigrum) and Zingiberaceae families (Turmeric, Ginger).
Most ovules have a funicle connected to the main body of the ovule for a short distance on the lateral side to create a ridge-like structure called Raphe.
The funiculus contains vascular tissues that transport food from the placenta to the ovule's body.
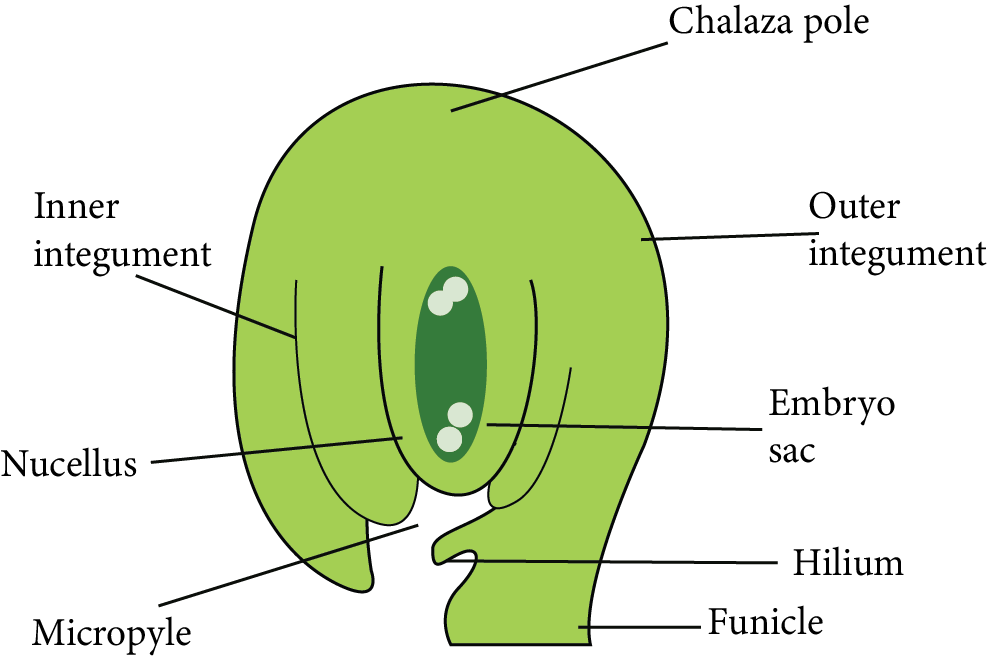
Structure of Ovule or Megasporangium
Types of Ovules:
Based On the Number of Integuments Present in Them:
Unitegmic: Ovules covered with a single integument are called unitegmic ovules. Examples include plants belonging to the Gamopetalae family and some of the gymnosperms.
Bitegmic: Ovules covered with two integuments are called bitegmic ovules. For example, Most Angiosperms.
Ategmic: Ovules that lack integuments are called ategmic ovules, like Santalum.
Based on the relative position of the micropyle, chalaza, and hilum with respect to the ovule’s body and funiculus orientation:
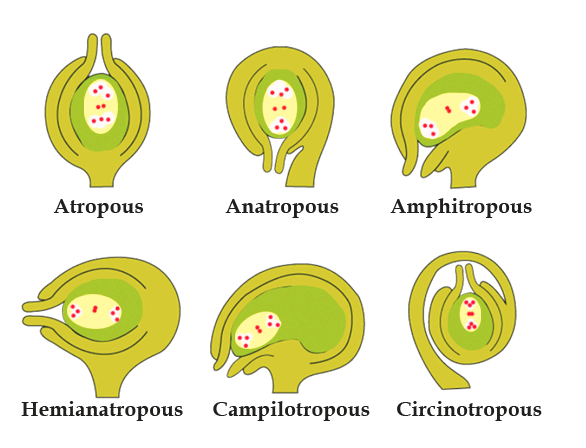
Types of Ovules
Orthotropous/ Atropous: Examples - Gymnosperms, Betel, Piper, etc.
Anatropous: Examples - Plant families like Compositae, Solanaceae, Cucurbitaceae, etc.
Hemitropous: Examples - Primula, Ranunculus.
Campylotropous: Examples - Legumes, Crucifers.
Amphitropous: Examples - Poppy, Mirabilis.
Circinotropous: Example - Opuntia.
Megasporogenesis:
Megasporogenesis is the process of forming a megaspore from a megaspore mother cell through meiotic division.
At the start of this mechanism, a single nucellus hypodermal cell differentiates and grows in size.
The presence of a distinct nucleus distinguishes it from the remaining cells. This special cell is known as an archesporial cell.
Archesporium divides the periclinally into two cells: a primary parietal cell (Outer side) and a primary sporogenous cell (Inner cell).
The primary sporogenous cell functions as a megaspore mother cell directly. It is diploid and meiotically divides to produce four haploid megaspores.
The four haploid megaspores are organised in a linear tetrad. The lowermost or the megaspore present at the chalazal end generally remains functional.
Out of the megaspore tetrad, the other three cells degenerate which are found towards the micropyle.
This functional megaspore gives rise to female gametophytes.
Development of Female Gametophyte (Embryo Sac):
Megagametogenesis: The process of formation of the embryo sac is defined as megagametogenesis.
The megaspore is the female gametophyte's first cell.
This megaspore expands in size and feeds on the nucellus. Megaspore nuclei divide mitotically to create two nuclei. Each nucleus shifts towards the opposite pole of the cell and arrives at its new location. The nuclei found at the poles split twice mitotically. As a result, four-four nuclei form at each pole. Thus, a total of eight nuclei are found in the megaspore at this stage.
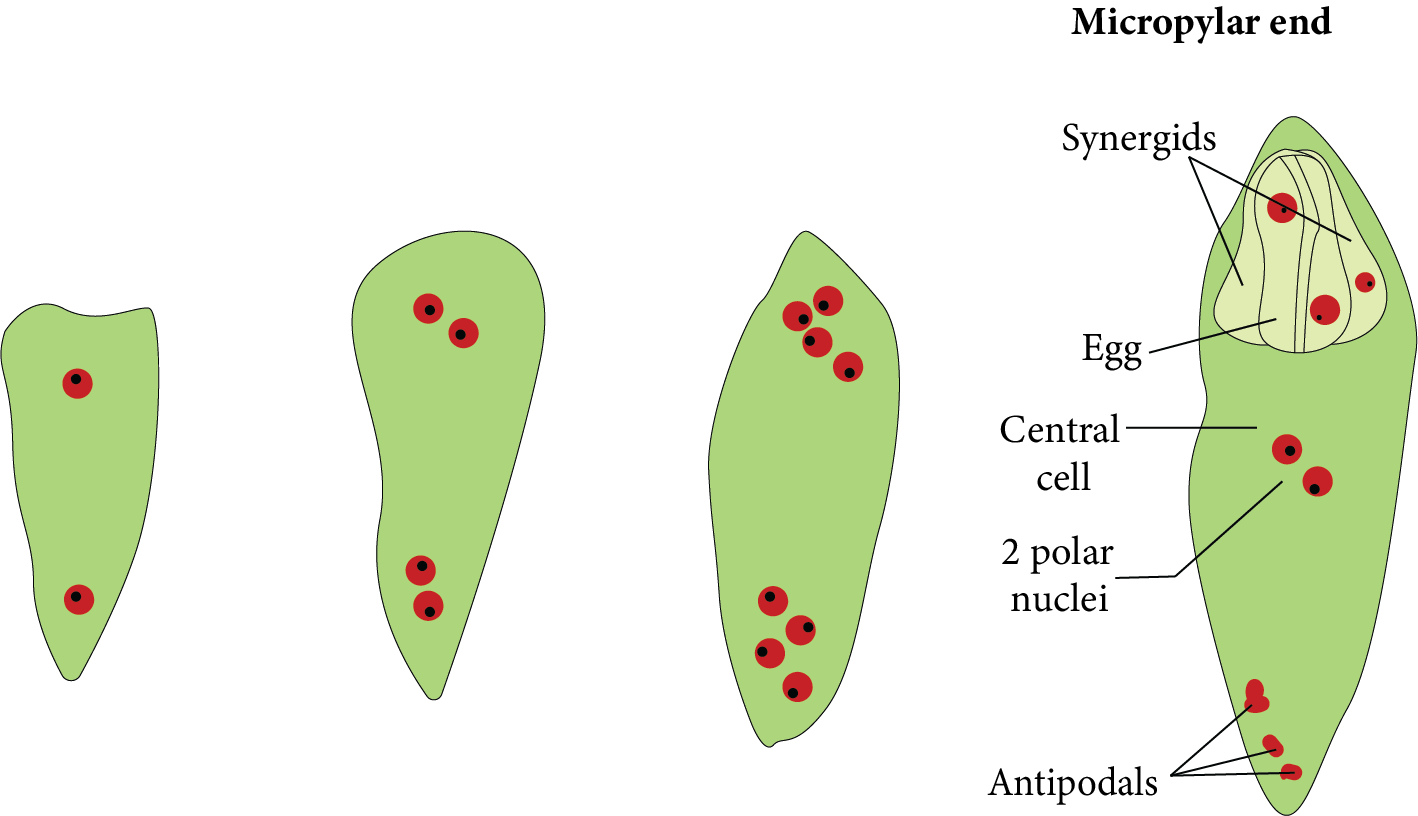
Development of Female Gametophyte
One nucleus from both poles [one from the chalazal and the other from the micropylar end] migrates towards the centre. They are referred to as polar nuclei and are present in the centre of the megaspore.
As a result of cytokinesis, the remaining three-three nuclei at both poles are encircled by cytoplasm to produce cells.
Three cells form towards the micropyle, one of which is larger and more distinct than the others. This is known as an egg cell, and the two remaining smaller cells are defined as synergids. The egg apparatus is made up of three micropylar cells. comprising one egg cell plus two synergids.
Antipodal cells are the three cells that form toward the chalaza. The central cell contains both polar nuclei. However, just before fertilisation, they merge or fuse with each other in the centre to produce the secondary nucleus. It is naturally diploid [2n].
As a result, a structure with seven cells and eight nuclei is formed, which is known as the female gametophyte or embryo sac of angiosperms.
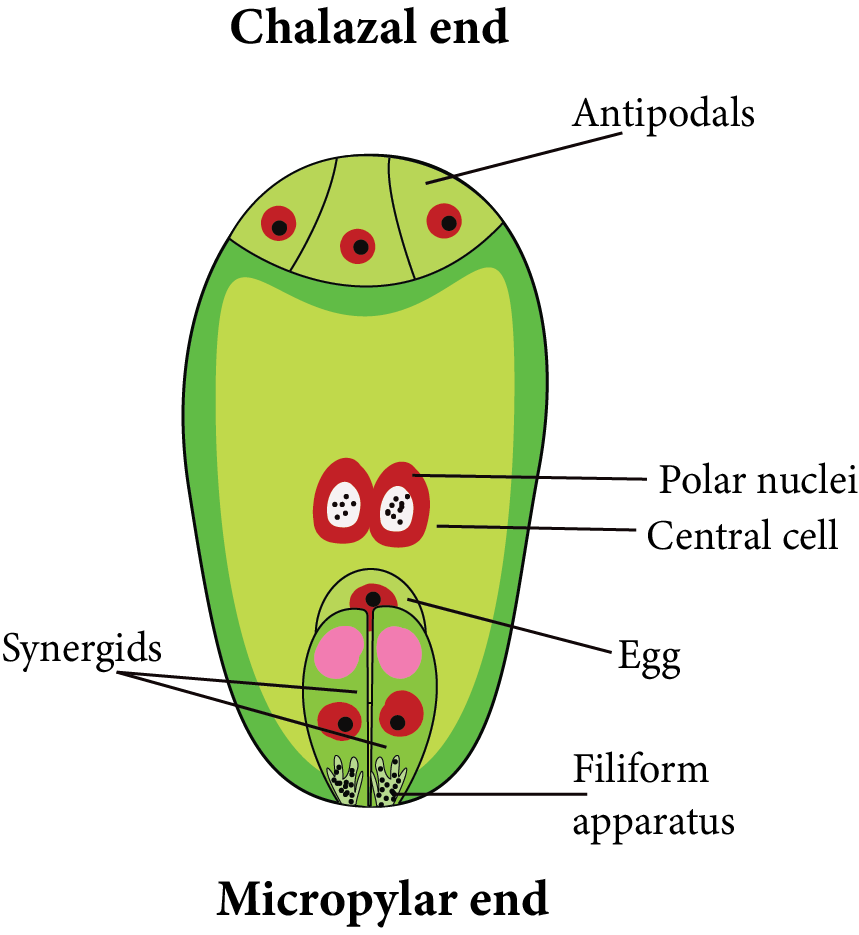
Female Gametophyte
Pollination
Pollination is the transport of pollen grains from the anther (male reproductive part) to the stigma (female reproductive part) of the same or different flowers of the same species.
Pollination is only found in seed-bearing plants which are gymnosperms and angiosperms.
There are two types of pollination: self-pollination and cross-pollination.
Self- pollination:
Self-pollination or autogamy occurs when pollen grains are transmitted from an anther to the stigma of the same or different flower, but of the same plant.
Adaptations for Self-pollinating Flowers:
Monocliny: Flowers are bisexual if they strictly follow self-pollination.
Homogamy: It occurs when both sex organs of a flower reach maturity at the same time. Mirabilis, potato, wheat, rice, and so on.
Cleistogamy: Some plants produce bisexual flowers that never open throughout their lives and are defined as cleistogamous flowers, Commelina and Viola, are examples of such flowers.
There are two types of flowers on the Commelina plant. Cleistogamous (closed) flowers are one type of flower, and chasmogamous (open) flowers are another.
Cross-pollination:
Cross-pollination or allogamy occurs when pollen grains are transmitted to the stigma of another flower of the same species. It occurs between two separate flowers.
There are two types of cross-pollination: geitonogamy and xenogamy.
Geitonogamy: Geitonogamy occurs when pollination occurs between two flowers of the same plant. It is self-pollination genetically since all flowers belonging to the same plant are genetically related. However, ecologically, it is regarded as cross-pollination.
Xenogamy: When pollination occurs between two different flowers belonging to two different plants of the same species, this is referred to as xenogamy. This is genuine cross-pollination. It is both genetically and ecologically significant.
Adaptations for Cross-pollinating Flowers:
Dicliny: The flower's unisexuality confirms cross-pollination. Self-pollination does not occur in these flowers. It means that allogamy is now required for cross-pollination. Date palms, cucumber, Cucurbita, and others are examples.
Dichogamy: It occurs when the stamens and carpels of a flower do not reach maturity at the same time in many hermaphrodite flowers of the plants. There are two kinds of dichogamy–
Protandry occurs when the anther of a flower matures before the carpels. It's the most common. For example Salvia.
Protogyny occurs when the flower's carpels mature before the stamens. It is found in a small number of plants, including Ficus, Saraca indica, and the majority of the Cruciferae and Rosaceae families.
Herkogamy: In some plants, morphological barriers develop between the anther and stigma of the same flower, preventing self-pollination as well as pollen grains from the anther from reaching the stigma of the same flower. Just cross-pollination is feasible in this condition. Gloriosa, Calotropis, and the Caryophyllaceae family are examples.
Heterostyly: Some plants' flowers have a difference in the length of the stamen filaments and the length of the style. As a result, self-pollination is not feasible in these plants, such as Primrose, Primula, etc.
Self-sterility/ self-incompatibility/ intraspecific incompatibility: In some plants, the pollen grains cannot actually grow on the stigma of the same flower in this condition. This is referred to as self-sterility. Petunia, Malva, Thea, Vitis, and Apple are examples.
Methods of Cross-pollination are:
Cross-pollination necessitates the transfer of pollen grains from one flower to another via external agents such as air, insects, water, birds, and so on.
Anemophily:
When pollen grains are relocated from one flower to another through the air, this is known as anemophily, and the flowers are known as anemophilous flowers.
Examples include all the gymnosperms, coconut, grasses, sugarcane, bamboo, and Cereal plants like rice, wheat, maize, etc.
Anemophilous flowers have the following features:
The pollen grains are produced by anemophilous plants in large amounts.
The pollen grains are small, light, and dry.
The stigma is hairy (brushy) and mucilaginous (Sticky).
Anemophilous flowers are neither beautiful nor fragrant.
They have no nectar glands.
Anemophilous flowers are almost always unisexual.
During wind pollination in Pinus, yellow clouds form in the sky due to its pollen grains and are known as "Sulphur Showers."
Entomophily:
Entomophily refers to pollination that occurs with the assistance of insects.
Honey bees are responsible for the majority of insect pollination (80%).
Only insects pollinate approximately 82 percent of angiosperms.
Flowers pollinated by insects are known as entomophilous flowers.
Entomophilous flowers have the following features:
The colour of these flowers is appealing.
They have a distinct scent.
There are also nectar glands.
Because of the presence of pollen kitt, the pollen grains of insect-pollinated flowers become sticky.
The stigma of flowers has a rough surface.
Nymphaea, Nelumbo, and Alisma are also hydrophytes and entomophilous plants.
Hydrophily:
It occurs when pollination is brought about by water.
It is classified into two types: Epihydrophily and Hypohydrophily.
Epihydrophily: When pollination occurs on the water's surface, as in Vallisneria and Ruppia.
Hypohydrophily: When pollination occurs within the water, as in Najas, Zostera, and Hydrilla.
Because pollen grains have specific gravity, they remain suspended in water.
Zoophily:
When animals pollinate plants, this is referred to as zoophily.
Flowers in zoophilous plants are typically large and attractive, with a high number of nectar glands.
Zoophily is classified into the following types:
Ornithophily: Ornithophily refers to the process of pollination by birds. For example, a sunbird or a hummingbird in a Begonia plant.
Chiropterophily: When pollination is carried out by bats, this is referred to as chiropterophily. Pteropus, for example. These flowers are huge in size like Anthocephalus.
Myrmecophily: It is the pollination by Termites and Ants. Prosopis kikar, Acacia, and so on.
Malacophily (Malmacophily): Snail pollination is known as malacophily. Orchids are a good example.
Ophiophily: Pollination by snakes is known as ophiophily. It occurs in Arisaema (Cobra Plant).
Pollen-Pistil Interaction:
It is a productive process that begins with pollen recognition and ends with pollen promotion or inhibition.
Pollen pistil interaction in plants provides a unique opportunity to elucidate the molecular basis of cell-cell communication as well as the evolution of intraspecific and interspecific barriers to fertilisation.
Emasculation and Bagging:
Emasculation is the process of removing stamens from flowers before they shed pollen to prevent self-pollination and allow cross-pollination.
To prevent unwanted pollen from contaminating the stigma of an emasculated flower, it must be covered with a suitable size bag, typically made of butter paper. This is known as bagging.
Fertilisation:
Fertilisation is the union/ merging or fusion of a male and a female gamete.
The fertilisation process is completed in four steps:
Pollen grain and pollen tube germination
Pollen tube entry into the ovule
Pollen tube enters the embryo sac.
Fusion of gametes
Pollen Grain and Pollen Tube Germination:
Pollen grains begin to grow on the stigma of the carpel following pollination.
They swell as they soak up moisture and sugar content from the stigma.
The intine layer of pollen grain expands out through any germinal pore of exine in the form of a tube-like outgrowth known as a pollen tube.
When the pollen tube descends from the stigma into the style, the vegetative nucleus is the first to enter the pollen tube, followed by the generative cell.
In pollen tubes, the tube nucleus is always at the end.
The vegetative nucleus regulates the growth of the pollen tube.
Meanwhile, the generative cell divides mitotically to create two male gametes.
Pollen Tube Entry Into the Ovule:
When the ovule reaches maturity, the pollen tube enters the ovary.
Obturators inside the ovary direct pollen tube passage to the micropyle.
A mature ovule, in which the embryo sac has also matured, has three paths for pollen tube entry:
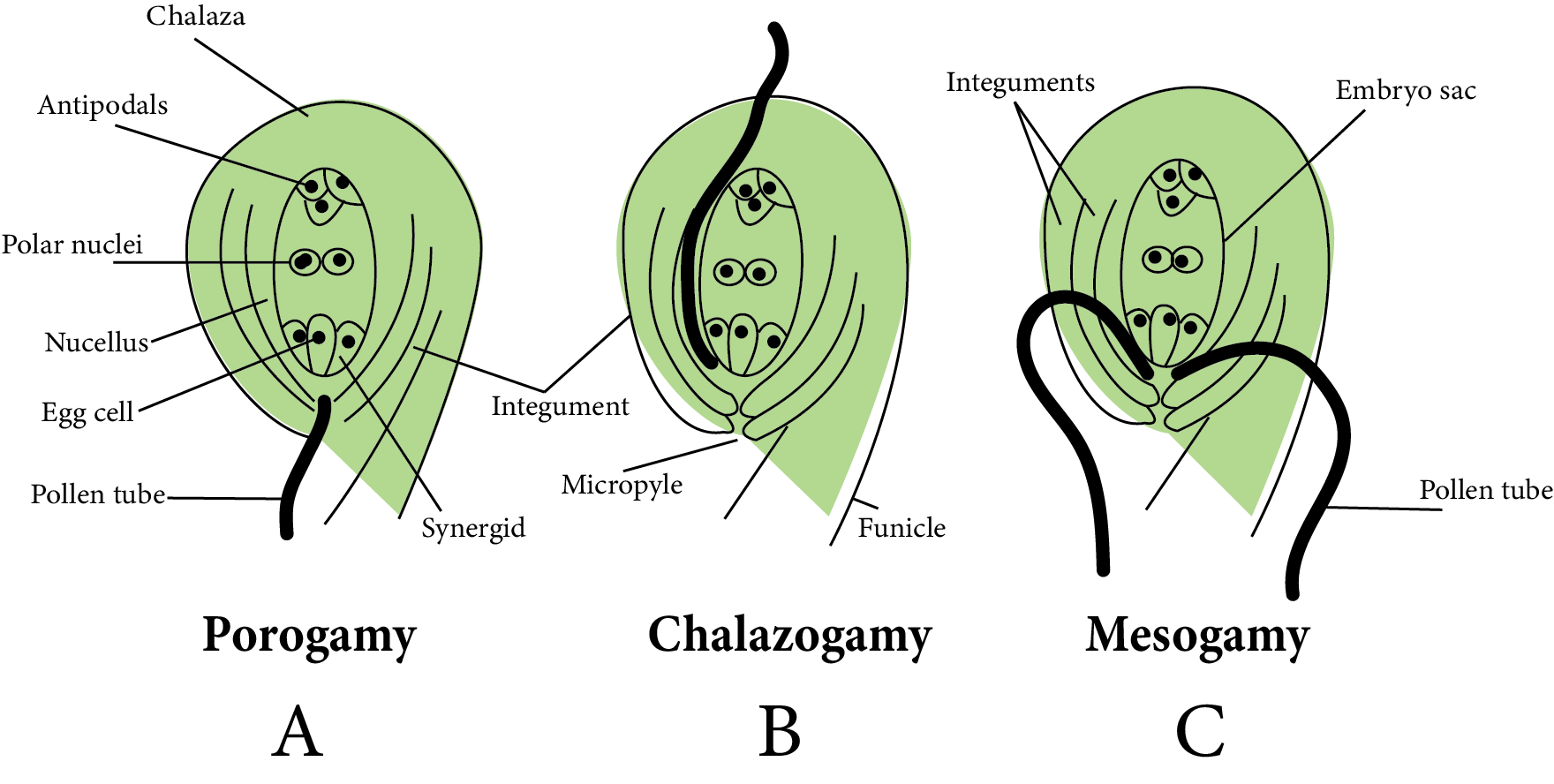
Pollen Tube Entry into The Ovule
Porogamy: It occurs when the pollen tube enters the ovule via the micropyle. It is found in the majority of Angiosperms.
Chalazogamy: The pollen tube enters the ovule via the chalaza in this method. As an example, consider walnuts.
Mesogamy: In this method, pollen tubes enter the ovule via integuments (for example, Cucurbita) or the funiculus (Example - Pistacia and Populus).
Pollen Tube Enters the Embryo sac:
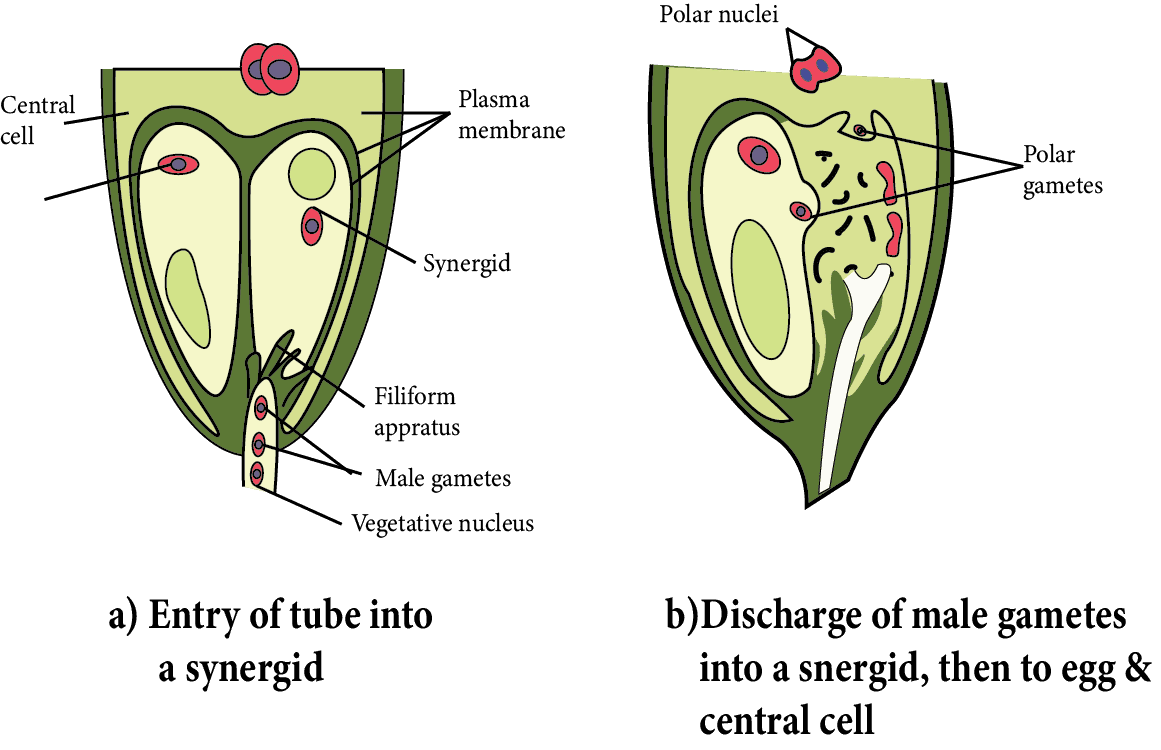
Pollen Tube Enters the Embryo sac
Inside the ovule, the entry of the pollen tube can be through any passage, but it can only enter the embryo sac through the egg apparatus.
Because synergid cells secrete a chemical that attracts the growth of pollen tubes, so, it grows towards the egg apparatus after entering the ovule. It means that the pollen tube moves chemotropically in the ovule.
When the pollen tube comes into contact with the egg apparatus, any synergid begins to degenerate.
Through the degenerating synergids, the pollen tube enters the embryo sac.
When the pollen tube tip enters the embryo sac, the vegetative nucleus degenerates.
After entering the embryo sac, the tip of the pollen tube swells and bursts.
Inside the central cell of the embryo sac, all contents of the pollen tube, including both male gametes, are released.
Fusion of Gametes:
The two polar nuclei of the central cell merged together to produce a diploid nucleus before or after the pollen tube entered the embryo sac. It is referred to as the secondary nucleus or the definitive nucleus.
One male gamete was fertilised with an egg cell and produced a diploid zygote. This fusion is referred to as syngamy. This is the actual mechanism of the fertilisation process.
The second male gamete merged with a diploid secondary nucleus formed by the merger of two polar nuclei. This is known as triple fusion, and it results in the formation of a triploid (3n) structure. It is known as the primary endosperm nucleus.
In angiosperms, fertilisation occurs twice at the same time, which is known as double fertilisation.
The universal feature of Angiosperm is double fertilisation and triple fusion. In double fertilisation, there are five nuclei and three gametes.
True fertilisation (syngamy) produces a zygote, which grows into an embryo.
By triple fusion, a triploid primary endosperm nucleus (PEN) is formed, which develops into endosperm, which serves as nutrition for the growing embryo.
After fertilisation, all of the remaining cells of the embryo sac, including antipodal cells and synergids, degenerate, with the exception of the zygote and primary endosperm nucleus. At this point, the zygote feeds on degenerating synergids and antipodal cells.
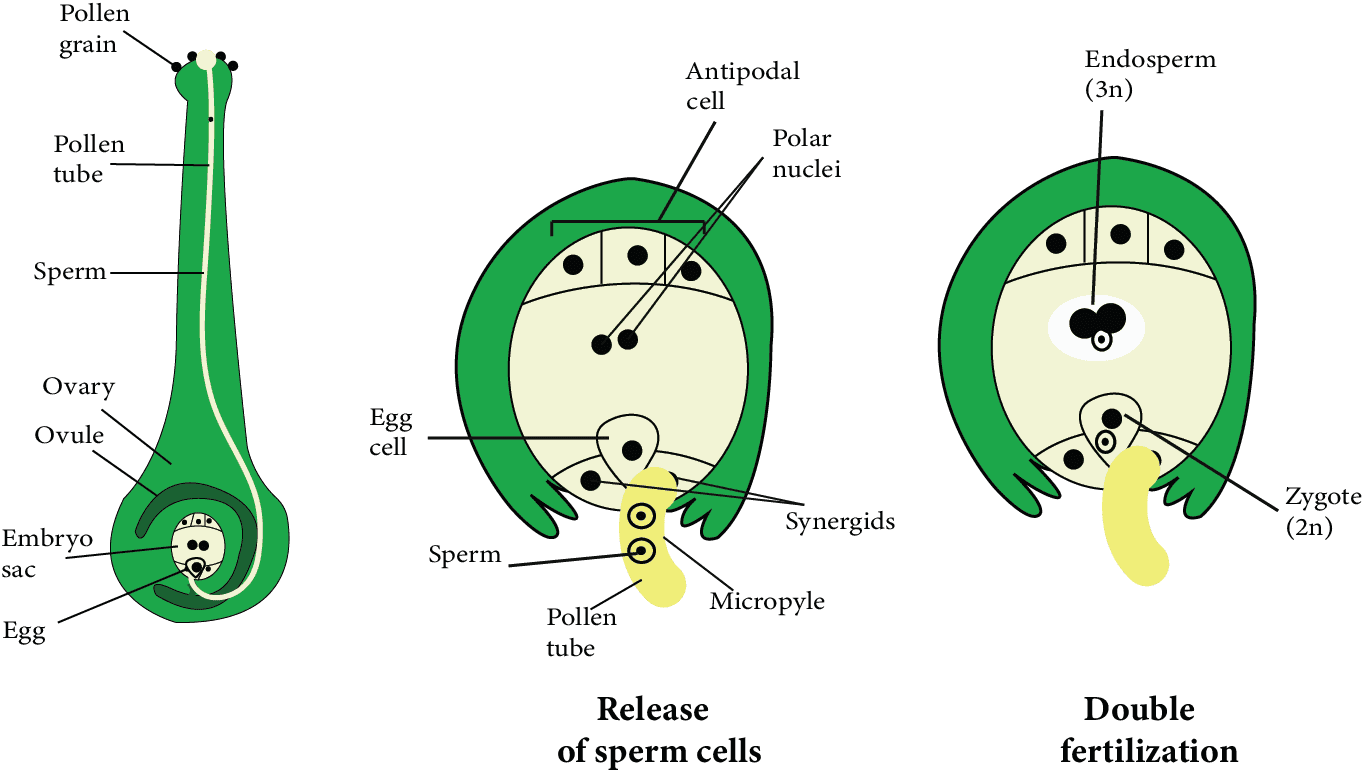
Fusion of Gametes
Endosperm Development
After fertilisation, the primary endosperm nucleus develops into endosperm, which stores food materials.
This food material is used by the embryo during the early stages of development and then again during seed germination. The endosperm contains food in starch form.
The endosperm is a nutritious tissue found in seed plants that feeds the embryo. It is almost always triploid.
It is created before fertilisation in Gymnosperms and is always haploid.
Types of Endosperm:
The endosperm is classified into three types based on development: nuclear, cellular, and helobial.
Nuclear Endosperm:
A nuclear endosperm is formed by free nuclear divisions of the nucleus of the primary endosperm, giving rise to a multinucleated endosperm.
Later, cytokinesis occurs, resulting in the formation of the multicellular endosperm.
It is the most popular Angiosperm endosperm (mostly occurs in Dicots).
The milky fluid present in green coconut is a form of nuclear endosperm known as a liquid syncytium.
Cellular Endosperm:
Each division of the primary endosperm nucleus (PEN) is followed by cytokinesis during development, ensuring that the endosperm remains cellular from the start.
This endosperm occurs in the Gamopetalae family, which includes Petunia and Datura.
Helobial Endosperm:
During the creation of this type of endosperm, the first division of the primary endosperm nucleus is pursued by unequal cytokinesis, resulting in the formation of two unequal-sized cells.
Each cell now undergoes free nuclear divisions and becomes multinucleated.
Cytokinesis eventually occurs at a time when it is transformed into a cellular endosperm.
Monocots have this type of endosperm. It is a type of intermediate endosperm.
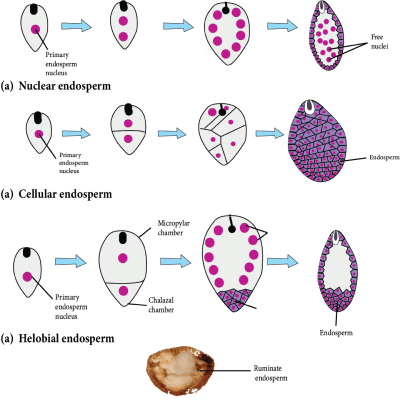
Types of Endosperm
Embryo Development
Embryogeny is the process of developing a mature embryo from a zygote or oospore.
Along with the endosperm development, the zygote begins dividing to produce an embryo.
Embryo Development in Dicots:
In angiosperms, the zygote goes through a resting phase. When the endosperm is formed, zygote development begins. It absorbs food from the endosperm at first and grows in size, then secretes a layer on its own later. It is now known as oospore.
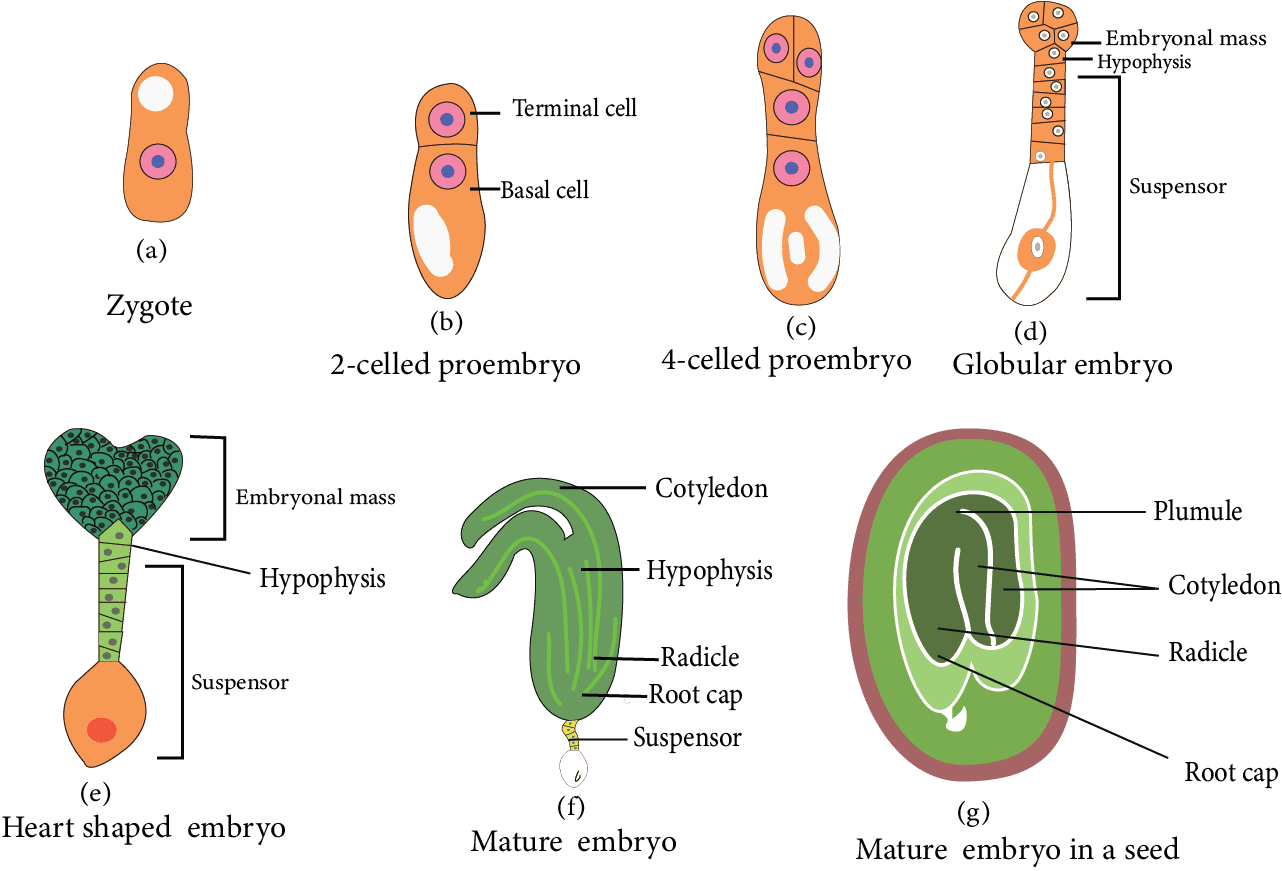
Embryo Development in Dicots
The oospore's first division is transverse. Two cells are formed as a result. The one cell closest to the micropyle is known as the basal cell or suspensor cell. The other cell, which forms towards the chalaza, is known as an apical cell, terminal cell, or embryonal cell.
Both the basal cell and the embryonal cell divide at the same time.
The basal cell divides transversely, while the apical cell divides vertically, yielding two suspensor cells and two embryonal cells.
In this stage, the embryo is made up of four cells that are arranged in a 'T' shape. To form four embryonal cells, embryonal cells divide vertically. This is the embryo's quadrant stage.
The two suspensor cells divide transversely, resulting in a 6-10 celled long filament-like structure known as a suspensor. Suspensor's primary function is to push the growing embryo into the food-rich endosperm to get nutrition.
The suspensor's micropylar cell swells. The haustorial cell is a type of suspensor cell.
The suspensor cell is located near the embryonal cells and is known as the hypophysis. This cell joins with the radicle to form the root's apex (Root cap).
These four-celled quadrant embryos divide transversely to generate an eight-celled embryo stage (called the octant stage). The octant's eight cells are divided into two tiers.
The four cells of the octant embryo located near the hypophysis are known as hypobasal cells, and the four cells located near the chalaza are known as epibasal cells.
The hypobasal cells give rise to the embryo's radicle and hypocotyl, while the epibasal cells give rise to the embryo's two cotyledons and plumule.
All of the cells in the octant divide periclinal, resulting in a 16-celled globular embryo (proembryo).
A heart-shaped embryo is formed as a result of the rapid division of embryonal cells in a globular embryo. This embryo's cells are all meristematic.
They create two cotyledons due to the rapid growth of two lobes of a heart-shaped embryo. Because of the curved position of the ovule's body, both growing cotyledons turn downwards.
Tissues can be found beneath the joining point of both. The cotyledons are capable of the formation of plumule, which is followed by the formation of epicotyl.
Radicles are formed by the tissues located opposite the plumule near the hypophysis. The hypophysis is responsible for the formation of the root cap. The torpedo or chordate stage refers to the embryo's curved position.
The embryonal axis exists between the plumule and the radicle.
In the Dicotyledon embryo, both cotyledons are visible at the lateral position of the embryonal axis, and the plumule is formed at the terminal position.
When an embryo develops, the suspensor dries and degenerates, resulting in meroblastic development.
The ovule has been modified into a seed in which the outer integument forms the testa and the inner integument forms the tegmen.
Only the ovule's micropyle remains unchanged and is also present in the seed.
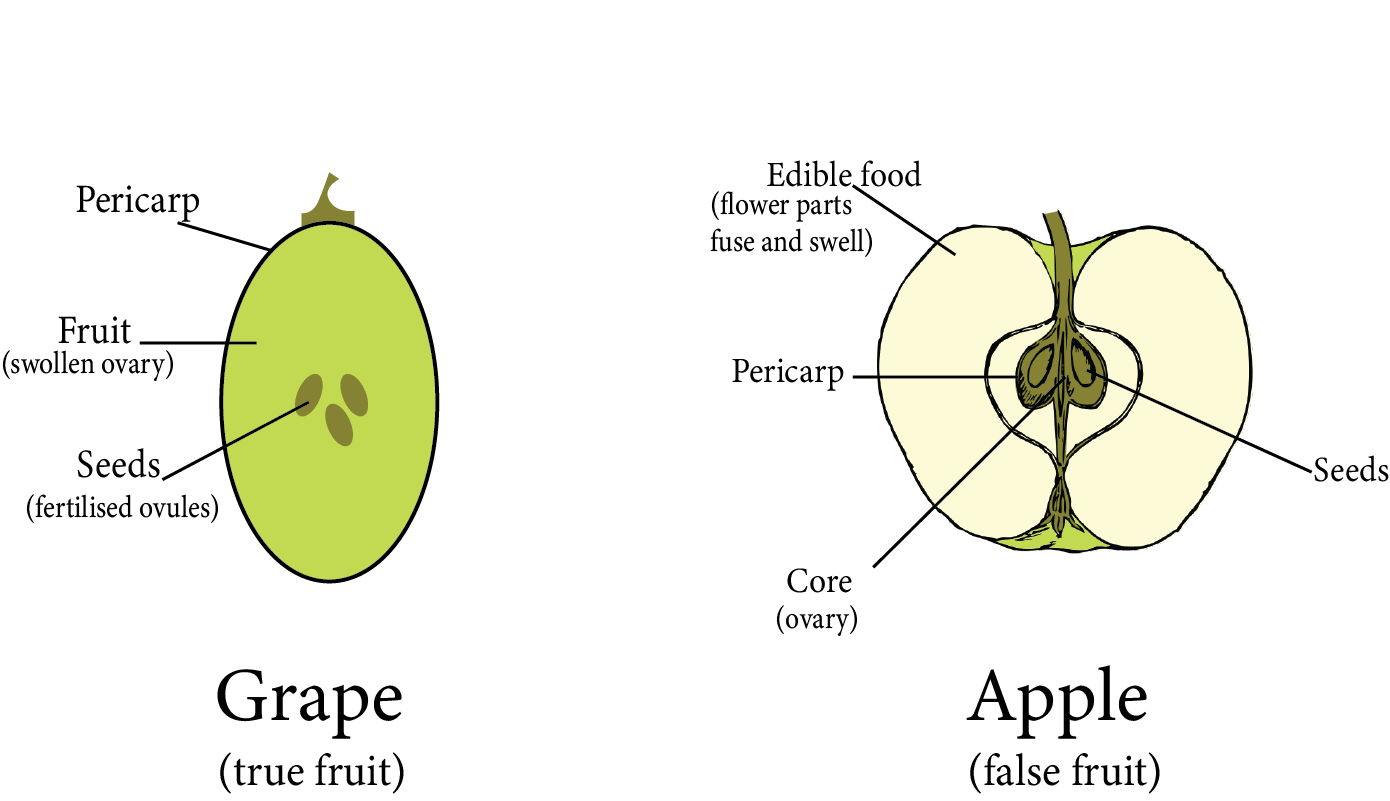
The entire ovary is transformed into fruit. This fruit is called true fruit because it is formed by a fertilised ovary.
Parthenocarpic fruit is produced by some angiosperms without fertilisation from the ovary.
Embryo Development in Monocots:
In the Oospore of monocot, the first division is the transverse division. As a result, two cells are formed: the upper chalazal cell is known as an embryonal cell, and the lower micropylar cell is known as a basal cell.
The basal cell does not split further but instead grows in size to form a single-celled vesicular suspensor.
Only embryonal cells divide transversely, with the terminal cell referred to as the cotyledon cell and the lower (middle) cell referred to as the embryonal axis cell.
A transverse division occurs in the embryonal axis cell, resulting in the formation of two cells.
One cell out of two produces plumule initial and another produces radicle initial.
The plumule initial cell divides to form an embryonic plumule whereas the radicle initial divides to form a radicle. Both initials are responsible for forming the embryo in a lateral position in this case.
Consistent divisions of cotyledon cells form an apical cotyledon. The scutellum is the name given to the apical cotyledon.
The plumule and radicle are protected by cap-like hard structures. The plumule covering is known as coleoptile, and the radicle covering is known as coleorhiza.
During the germination process, these structures are broken. In monocotyledons, the plumule is located laterally and the cotyledon is located terminally.
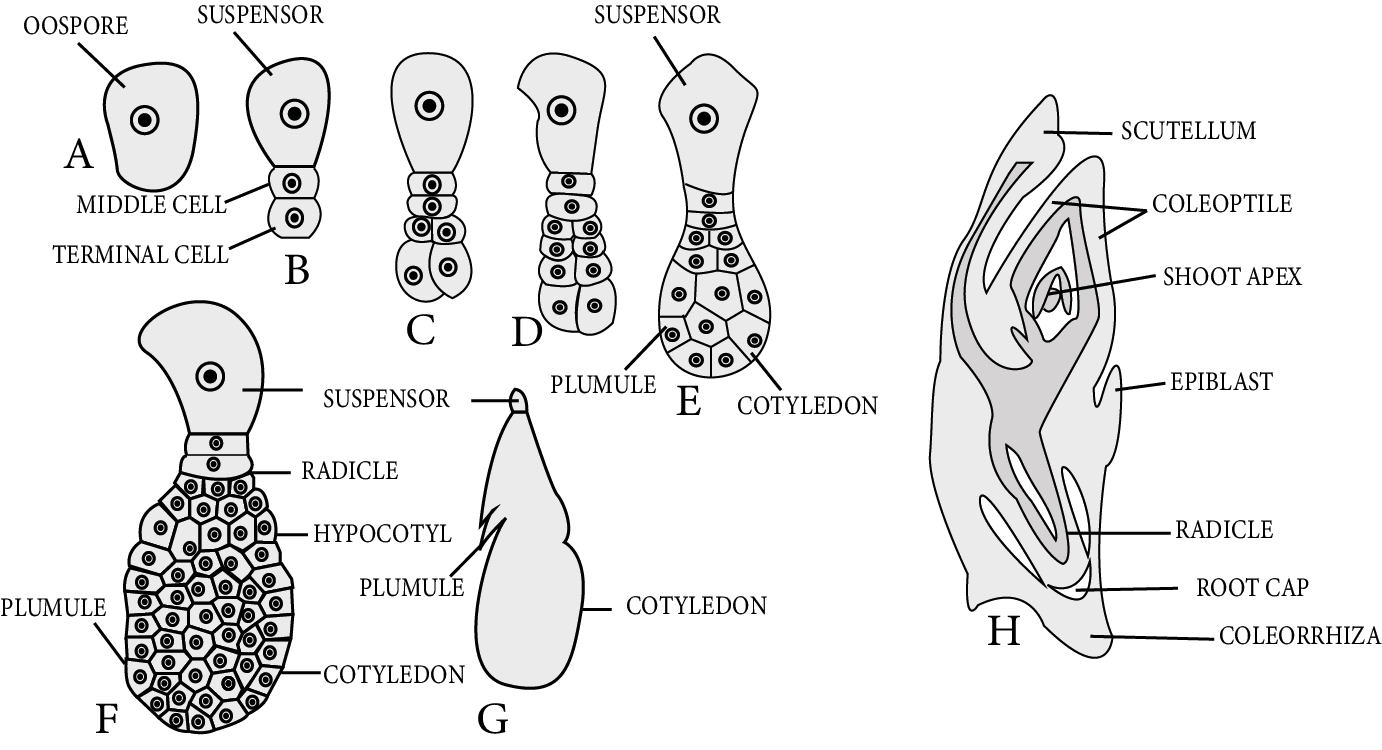
Embryo Development in Monocots
Structure of Seed:
A seed contains the cells which divide and develop to form an entire plant. After fertilisation, egg cells transform into embryos, ovaries transform into fruits, and ovules transform into seeds in angiosperms.
A ripened ovule is referred to as a seed morphologically. To put it another way, the seed is a mature, integument megasporangium.
Spermatophytes are distinguished by the presence of seeds (gymnosperms and angiosperms).
A seed is a dormant structure that contains protective coverings, reserve foods, and an embryo (2n).
The seed coat is the seed's outer protective covering, which develops from ovule integuments.
There are two distinct layers in the seed coat that develop from bitegmic ovules:
The outer layer is thick, hard, and leathery called testa developed from the outer integument.
The inner layer is thin and papery or membranous called tegmen developed from the inner integument.
There is a single-layer seed coat in seeds that develop from unitegmic ovules.
The seed is connected to the fruit wall or pericarp via a stalk known as a funicle or funiculus.
The hilum is the point at which the funiculus attaches to the mature seed body.
Water enters the seed through a small pore called the micropyle, which is located just near the hilum.
Because most angiosperm ovules are inverted or anatropous, a ridge known as the raphe forms.
Depending on whether the seed is monocot or dicot, the embryo has an embryonal axis called tigellum to which one or two cotyledons are attached.
The portion of the tigellum below the cotyledon attachment point is known as the hypocotyl, and it bears a radicle (future root) at its tip. Similarly, epicotyl refers to the portion of the tigellum above the attachment site of the cotyledons that bears a plumule (future shoot) at its tip.

Structure of Seed
The embryo's cotyledons are simple structures that are generally thick and swollen due to the storage of food reserves, as in legumes, while in cereals, there is a special nourishing tissue called endosperm.
Perisperm refers to the unused portion of the nucellus that is present as a thin layer around the endosperm. For example, black pepper, castor, and so on.
The scutellum is a solitary cotyledon found in Monocot embryos.
Types of seeds:
Endospermic or Albuminous:
Food is deposited in the endosperm of these seeds. These seeds are known as endospermic seeds or albuminous seeds.
Examples of endospermic dicot seeds are Castor, Papaya, Cotton, etc.
Non-endospermic or exalbuminous:
Because these seeds lack an endosperm at maturity, they are referred to as non-endospermic.
Examples of non-endospermic dicot seeds are Gram, Bean, Pea, Cucumber, Tamarind, etc
Germination of Seeds:
Germination is the initiation of seedlings from a dormant embryo of a seed after it resumes active growth.
In germinating seeds, the rate of respiration increases.
Germination is classified into two types based on the behaviour of the cotyledons: epigeal and hypogeal.
Epigeal germination: Cotyledons are pushed out of the soil as a result of hypocotyl growth or elongation. This is known as epigeal germination. For example, beans.
Hypogeal germination: Because of the first epicotyl elongation, plumule emerges from the ground while cotyledons remain underground. This is known as hypogeal germination. For example, gram.
Dormancy of Seeds:
The presence of dormancy in a seed is its most distinguishing feature. Because of this quality, seeds can live for many years. The seeds travel long distances via water, air, or insects.
Most seeds do not germinate immediately after dispersal. They eventually germinate.
The period of time between seed maturation and germination is known as the "dormancy period."
Formation of Fruit:
Fruit is a fertilised ovary with or without a seed, as well as any accessory structures that are closely associated with it.
True fruits are those that develop solely from the ovary and do not involve any other floral parts in their development. For example, grapes.
False fruits are fruits in which, in addition to the ovary, other floral parts contribute to their development. For example, aggregate fruits.
Seedless fruit or parthenocarpic fruits are those that develop without fertilisation and thus lack seeds, such as bananas, guavas, and apples.
Types of Fruits based on the Parts of the Flower Participates in Fruit Formation
Apomixis, Parthenocarpy, Polyembryony
Apomixis is a type of asexual reproduction wherein seeds form without gamete fusion. It includes the process by which a diploid nucellus cell grows into an embryo, producing a diploid seed with the same genetic makeup as the parent.
Parthenocarpy occurs when the fruit develops in an unfertilized flower, actually resulting in a seedless fruit. It can occur naturally in pineapple, grapes, apple, pear, and banana varieties.
Polyembryony is the existence of multiple embryos in a seed. It could be because there is more than one egg cell in the embryo sac or there can be more than one embryo sac in the ovule, and all of the egg cells may be fertilised.
Key Points to Remember
In angiosperms, flowers are the site of sexual reproduction.
The male reproductive organs are represented by stamens in the flower, while the female reproductive parts are represented by pistils.
An anther, male reproductive part, is typically bilobed, dithecous, and tetrasporangiate. Pollen grains are formed inside the microsporangia.
The microsporangium is surrounded by the four wall layers epidermis, endothecium, middle layers, and tapetum.
The sporogenous tissue cells in the microsporangium exhibit meiosis to form microspore tetrads. The process is known as microsporogenesis.
Pollen grains develop from individual microspores. The male gametophytic generation is represented by pollen grains.
The pollen grains are two-layered, the outer layer called exine and inner layer called intine. The exine is composed of sporopollenin and contains germ pores.
Pollen grains can have two cells, one vegetative or tube cell that forms a pollen tube and another is a generative cell that splits into two male gametes.
The stigma, style, and ovary are the three parts of the pistil. The ovary contains ovules. The ovules have a funicle stalk, protective integuments, and a micropyle pore. The nucellus is the central tissue wherein the archesporium develops.
The megaspore mother cell, an archesporium cell, divides meiotically, and one of the megaspores produces the embryo sac which is the female gametophyte. The mature embryo sac has seven cells and eight nuclei. The egg apparatus, which consists of two synergids and an egg cell, is located at the micropylar end. Three antipodals are located at the chalazal end. A large central cell with two polar nuclei is located in the centre.
Pollination is the process by which pollen grains are transferred from the anther to the stigma.
Pollination is the process by which pollen grains are transferred from the anther to the stigma. Pollinating agents can be abiotic like wind and water or biotic like animals.
Pollen-pistil interaction encompasses all events from pollen grain landing on the stigma to pollen tube entry into the embryo sac (if pollen is compatible) or pollen inhibition (if pollen is incompatible).
So after compatible pollination, pollen grain grows on the stigma, and the resulting pollen tube grows through the style, into the ovules, and eventually releases two male gametes in one of the synergids.
Angiosperms have double fertilisation since two fusion events, syngamy and triple fusion, occur in each embryo sac. The diploid zygote as well as the triploid primary endosperm nucleus are the results of these fusions.
The embryo develops from the zygote, and the primary endosperm cell constitutes the endosperm tissue.
Endosperm formation always comes before embryo development.
Before maturation, the developing embryo goes through various stages such as proembryo, globular, and heart-shaped. An embryonal axis along with epicotyl and hypocotyl exists in mature dicotyledonous embryos.
Monocotyledon embryos have a single cotyledon.
Following fertilisation, the ovary grows into fruit and the ovules develop into seeds.
Apomixis is a phenomenon found in certain angiosperms, particularly grasses. It leads to formation of seeds in the absence of fertilisation. Apomicts offer numerous benefits in horticulture and agriculture.
Some angiosperms have multiple embryos in their seed. This is referred to as polyembryony.
Sexual Reproduction in Flowering Plants: Importance for NEET
In these revision notes, the chapter provides information on the process of sexual reproduction in flowering plants. The bisexual flowers have both male (stamens) and female (carpels or pistils) reproductive parts or organs. Whereas, either of the two reproductive organs is present in unisexual flowers, such as corn, in which the tassels have the male parts (stamens) and the ears or silk have the female parts (styles and stigma).
Stamen is bascially the male reproductive unit called Angiosperm. Anther, on the other hand, the bilobed structure having two thecae in each lobe. In the chapter, students are further explained about the Microsporogeneis, the formation of microspores from the pollen mother cell. While you study further, you will find that during the process of sexual reproduction, the pollen grains multiplies forming the male gametophytes. Students will also know about the development of female gametophyte in different stages.
Benefits of Sexual Reproduction of Flowering Plant Notes
The Sexual Reproduction in Flowering Plants notes for NEET are formulated by subject experts of Vedantu. Each topic of the chapter is well explained, making it easy to learn and understand for students. Phases of reproduction, like pre-fertilization, and post-fertilization, are explained in detail.
Throughout the chapter, students will learn the significance of fertilization in flowering plants.
Students can understand the concept well and prepare for the NEET through these notes. They can also use these notes for the last-minute revision.
Download Sexual Reproduction in Flowering Plants Notes for NEET PDF
You can now prepare well for the NEET exam by downloading sexual reproduction in flowering plants Class 12 notes PDF. You can refer to these PDF notes to understand the complete reproduction process and also to understand the process of development of female gametophyte. The best part of using the PDF file is you can do the last-minute revision as a part of NEET preparation.
NEET Biology Revision Notes -Chapter Pages
Other Important Links

FAQs on Sexual Reproduction in Flowering Plants Notes for NEET 2024
1. What exactly is the Sexual Reproduction in Flowering Plants?
Male and female sex organs can also be found in plants. Therefore, sexual reproduction in flowering plants is basically the process to produce new plants through the fusion of male and female gametes. In the process of sexual reproduction, a new seed is generated when the male gamete unites with the female gamete.
2. What is asexual reproduction in Flowering Plants?
In asexual reproduction process, new plants are born without production of seeds. It is a process in which new plants are produced from roots, stems, buds, and leaves. Since reproduction is through the vegetative parts of the plant, it is known as vegetative propagation.
3. What is Pollination in Sexual Reproduction of Flowering Plants?
Pollination is the mechanism that helps in the process of sexual reproduction in flowering plants. In other words, Pollination is the transfer of pollen grains towards the stigma of the pistil, which results in the fertilization of the flower and formation of the new seed.
4. What are the 3 types of Asexual Reproduction in plants?
The 3 types of Asexual reproduction includes- fragmentation, budding, and vegetative propagation.